Vekst analyse avhenger av et sett med verktøy som ofte brukes av planteforskere til å beskrive genotype bestemt vekstforskjeller og / eller fenotypiske responser på miljøfaktorer. De inkluderer størrelse og vekt målinger av hele anlegget eller et orgel og beregninger av vekstrater for å utforske de underliggende mekanismene for vekst. Organ vekst bestemmes av celledeling og ekspansjon på cellenivå. Derfor, inkludert kvantifisering av disse to prosessene i vekst analyser er nøkkelen til å forstå forskjeller i hel-organ vekst en. Følgelig er det avgjørende å ha en passende metode for å bestemme cellevekstparametre som er forholdsvis lett å bruke av ikke-spesialiserte laboratorier.
Er allerede etablert kinematisk analyse som en tilnærming som gir et kraftig rammeverk for utvikling av organvekstmodeller 2. Teknikken har blitt optimalisert for lineære systemer,slik som Arabidopsis thaliana røtter og enfrøbladede blader, men også for ikke-lineære systemer, slik som tofrøbladede blader 3. I dag er denne metodikken i økende grad blir brukt til å studere hvordan genetiske, hormonelle, utviklingsmessige, og miljøfaktorer påvirker celledeling og ekspansjon i ulike organer (tabell 1). Dessuten gir det også et rammeverk for å knytte cellulære prosesser til sine underliggende biokjemiske, molekylære og fysiologiske bestemmelser (Tabell 2), selv om begrensningene kan pålegges av orgel størrelse og romlige organiseringen for teknikker som krever større mengder plantemateriale (f.eks metabolitten målinger, proteomikk, etc.).
Enfrøbladede blader, som for eksempel mais (Zea mays) blad, representere lineære systemer i hvilke celler beveger seg fra undersiden av bladet mot spissen, sekvensielt passerer gjennom meristem og forlengelse sone for å oppnå den modnesone. Dette gjør det til et ideelt modellsystem for kvantitative studier av romlige mønstre av vekst fire. Videre mais blader har store vekstsonene (meristem og forlengelse sone som strekker seg over flere centimeter 5) og gi muligheter for studier på andre nivåer i organisasjonen. Dette gir mulighet for undersøkelse av (antatte) reguleringsmekanismer som styrer celledeling og vekst, kvantifisert ved kinematisk analyse gjennom en rekke molekylære teknikker, fysiologiske målinger og cellebiologi tilnærminger (tabell 2).
Her gir vi en protokoll for å utføre en kinematisk analyse i monocot blader. Først forklarer vi hvordan du kan gjennomføre en skikkelig analyse av både celledeling og celleforlengelse som en funksjon av posisjon langs bladet aksen og hvordan man regner kinematiske parametre. For det andre viser vi også hvordan denne kan brukes som et grunnlag for prøvetaking utforming. Her diskuterer vi to tilfeller: høyoppløselig sampling end fokusert prøvetaking, slik at bedre data tolkning og lagring av tid / penger, henholdsvis.
Tabell 1. Oversikt over kinematiske analyser fremgangsmåter for kvantifisering av celledeling og vekst i forskjellige organer.
| organ | henvisning |
| monocotyledonous blader | 16, 20, 21, 22 |
| root tips | 2, 23, 24, 25, 26, 27, 28, 29 |
| dicotyledonous blader | 21, 30, 31 |
| skyte apikale meristem | 32 |
Tabell 1. Oversikt over kinematiske analyser fremgangsmåter for kvantifisering av celledeling og vekst i forskjellige organer.
<p class="jove_content" fo:keep-together.within-page = "1">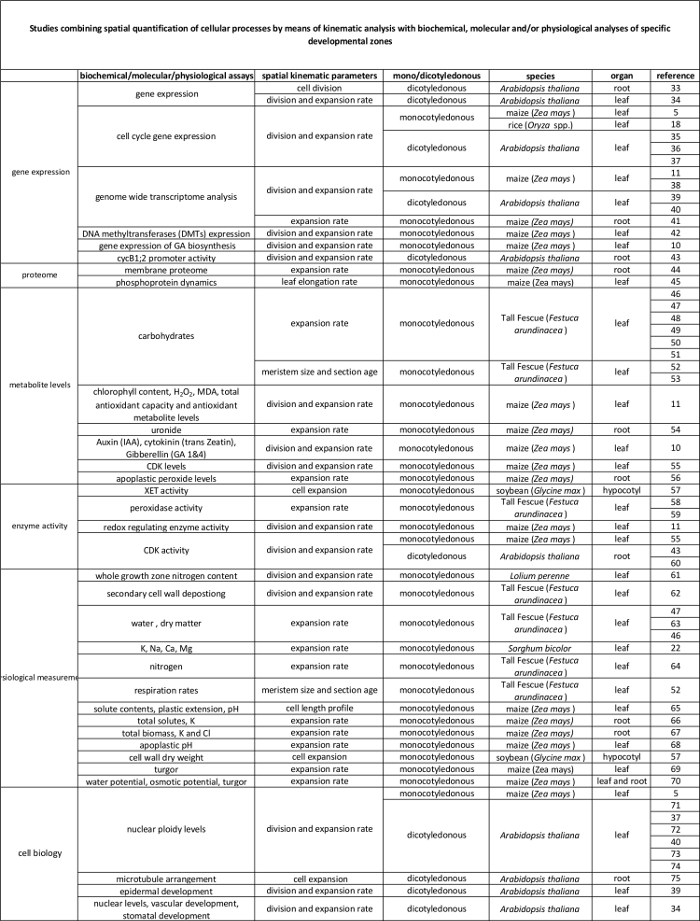
Tabell 2. Kobling mellom celleprosesser kvantifisert ved den kinematiske analysen til deres regulering på molekylnivå. Referanser til ulike studier som forbinder kvantifisering av cellulære prosesser til resultatene fra biokjemiske og molekylære assays i forskjellige arter og organer. Xyloglucan endotransglucosylase (XET), malondialdehyde (MDA), cyclin-avhengige kinaser (CDK). Klikk her for å se en større versjon av denne tabellen.
