In single cultures, both the hCMEC/D3 cells (Figure 3A) and the primary mouse BECs (Figure 3B) form meshwork-like structures throughout the well. The structures are characterized by a meshwork of interlinked nodes (Figure 3). In all the paradigms described (Figure 1), the meshwork-like structures are similar after 24 h in the control groups, forming ~20 meshwork-like structures with a total cellular length of ~10,000 pixels.
To highlight the applicability of the methods to disease relevant stressors, oAβ was added to the hCMEC/D3 and the primary mouse BECs in two paradigms. In the disruption of cell meshwork formation (Paradigm 1), oAβ and cells are plated at the same time. In the disruption of preformed meshwork (Paradigm 2), oAβ is added 4 h after plating the cells (see Figure 1A). oAβ at 100 nM induces disruption of the meshwork-like formation and degeneration of preformed meshwork (Figure 3A, 3B). For example, using the hCMEC/D3 cells in both paradigms, quantification of the total cell coverage/length is 16-20% lower with 100 nM oAβ17. Further, the number of meshes is reduced by 40% with 100 nM oAβ in both paradigms17. Thus, a disease relevant stressor induces a similar detrimental effect on human and mice BEC coverage.
A key advantage of the cell culture system is the ability to identify factors or treatments that prevent disease-relevant damage, which can then advance to in vivo testing. As a proof of principle experiment, the effects of the main angiogenic growth factors on preventing oAβ-induced changes to vessel coverage were assessed17. The EGF prevented oAβ-induced damage to the hCMEC/D3 cells. Based on those data, EGF was tested in a prevention paradigm using a transgenic mouse model that recapitulates critical aspects of AD-like pathology. EGF treatment prevented the cognitive and BBB deficits, including vessel degeneration23. These data support the predictive potential of the in vitro system for in vivo activity. Currently, we utilize all three developed paradigms for screening: 1) mesh formation, 2) prevention of cell meshwork disruption, and 3) simultaneous treatment of meshwork disruption. As highlighted in Figure 3, EGF can protect against oAβ-induced damage in immortalized and primary mouse BEC cultures.
The BBB consists of BECs, pericytes and astrocytes that collectively contribute to overall cerebrovascular coverage. Therefore, one adaptation of the in vitro system is the incorporation of all three BBB cell types. Our triple culture assay paradigm incorporates the hCMEC/D3 cells, primary human pericytes and primary human astrocytes, which are added sequentially to the basement membrane matrix (Figure 1B). In the triple culture assay paradigm, the hCMEC/D3 cells form a similar meshwork pattern as in the single cultures, but with the pericytes and astrocytes attached to the nodes and connecting branches (Figure 4). The addition of 100 nM oAβ induces meshwork disruption (~10-15%) and a reduction in the number of pericytes contacting the BECs17. In correlation with the data derived from the single culture assay paradigms, oAβ-induced damage is prevented by EGF treatment. Thus, the triple culture BBB model is well suited for studies focused on the interactive effects of astrocytes, pericytes and BECs on vessel dynamics.
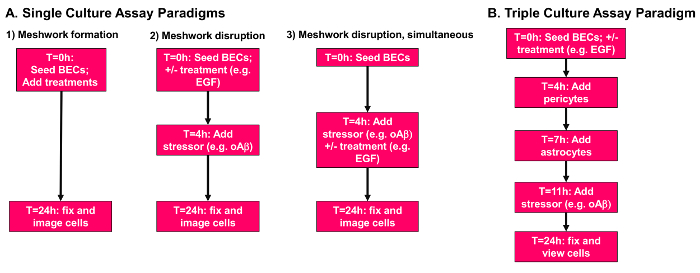
Figure 1: Overview of meshwork formation and disruption assays. (A) Single culture assay paradigms. Paradigm 1, meshwork formation. Cells, stressors and treatments are all added to the plate at the 0 h time point. Paradigm 2, prevention of meshwork disruption. Cells are plated in the presence of desired treatments, incubated for 4 h, and a stressor is added at the 4 h time point. Paradigm 3, simultaneous treatment of meshwork disruption. Cells are plated and allowed to form meshwork-like structures for 4 h before treatments and/or stressors are added simultaneously at the 4 h time point. All paradigms end at the 24 h time point, and cells are fixed with 4% paraformaldehyde. (B) Triple culture assay paradigm. BECs (10,000 cells/well) are plated in the presence of desired treatments at 0 h. At the 4 h time point pericytes (2,000 cells/well) are gently added to the plate. At 7 h astrocytes (10,000 cells/well) are added to the plate, followed by the addition of relevant stressors at 11 h. Cells are then incubated until the 24 h time point and fixed with 4% paraformaldehyde. Please click here to view a larger version of this figure.

Figure 2: Quantitative analysis. All images are opened on Fiji ImageJ and batch-processed using the Angiogenesis Analyzer22. Quantification of total length and number of meshes are utilized. Please click here to view a larger version of this figure.
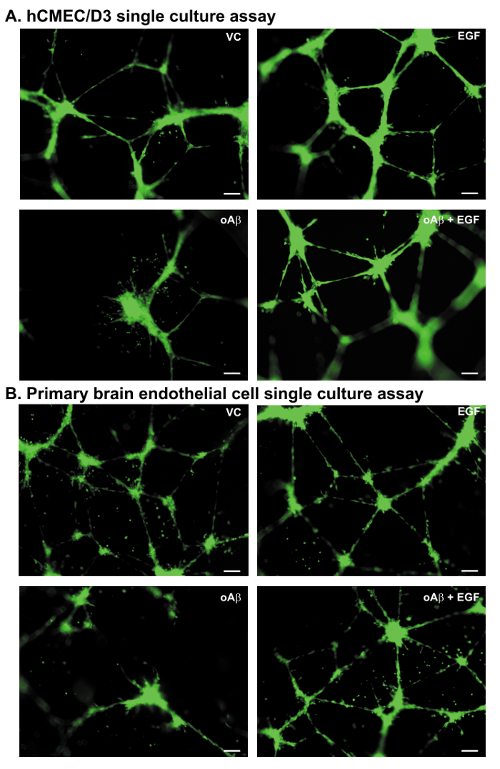
Figure 3: Meshwork disruption assays: representative images. All images are derived from experiments utilizing Paradigm 2, meshwork disruption. (A) Representative images of the hCMEC/D3 cells plated and treated with vehicle control (VC), EGF (100 nM), oAβ (100 nM), or oAβ (100 nM) + EGF (100 nM). Images at 10X magnification, Scale bar = 100 µm. (B) Representative images of the primary mouse BECs plated and treated with VC, EGF (100 nM), oAβ (100 nM) or oAβ (100 nM) + EGF (100 nM). Images at 10X magnification, green = BECs, Scale bar = 100 µm. Please click here to view a larger version of this figure.
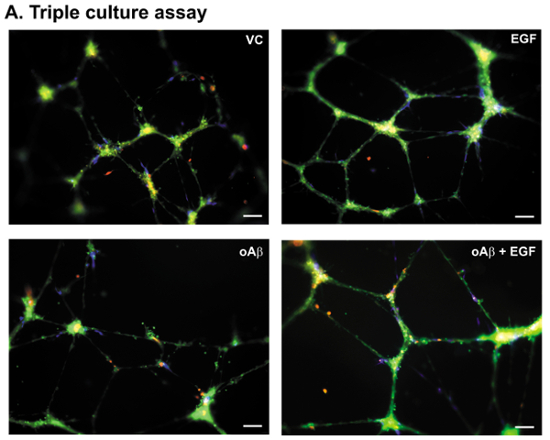
Figure 4. Triple culture assay: representative images. (A) Representative images of the hCMEC/D3 cells, primary human pericytes, and primary human astrocytes plated and treated with VC, EGF (100 nM), oAβ (100 nM), or oAβ (100 nM) + EGF (100 nM) according to the paradigm described in Figure 1B. Images at 10X magnification, green = BECs, blue = pericytes, and red (pseudocolor) = astrocytes. Scale bar = 100 µm. Please click here to view a larger version of this figure.
