Expression of total tau and phospho-tau was examined after treating the cells with different concentrations of curcumin or LiCl (Figure 1). Treatment of cells with the three different concentrations of curcumin decreased tau expression levels; however, phospho-tau expression increased upon treatment with low concentration of curcumin but decreased upon treating cells with higher curcumin concentrations. Anti-phospho-tau (Ser396) was used for detection of phospho-tau. Levels of both total tau and phospho-tau decreased upon treatment of the cells with the three different concentrations of LiCl (Figure 1). Previous research had shown that tau expression levels vary in different cancer types and at different sites of the same cancer types. Previous data on colorectal cancer showed that tau was expressed in two cell lines (HCT 116 and SW480) of colorectal cancer that were also phosphorylated19. Phospho-tau levels in cells treated with 5 µM curcumin were higher than those in untreated cells. Phospho-tau levels were higher than total tau in cells treated with the same concentration of curcumin. Treatment of cells with 10 µM curcumin decreased phospho-tau levels compared with untreated cells but phospho-tau expression was still more than total tau expression levels. Treatment of cells with 30 µM curcumin lowered phospho-tau expression compared with phospho-tau in untreated cells and total tau levels in the same treated cells.
This manuscript established an easy protocol for assessing the phosphorylation status of tau by a phosphatase assay (Figure 2). Following curcumin treatment, overall tau phosphorylation did not change significantly and phosphorylation at specific amino acid residues was significant (Figure 1). Overall tau phosphorylation was stopped in cells treated with LiCl as compared with untreated cells. Phosphatase-treated samples electrophoresed faster than untreated samples, verifying that untreated samples were more hyperphosphorylated than phosphatase-treated samples. Curcumin-treated cells showed nearly the same results, indicating that treatment of colorectal cancer cell lines did not reduce tau phosphorylation as shown in Figure 1. Both phosphatase-treated and untreated samples electrophoresed at nearly the same range in samples of cells treated with LiCl, indicating that tau phosphorylation in these cells was reduced (Figure 2). Here, high concentrations (20 µM or 30 µM) of curcumin-treated samples were taken to compare overall phosphorylation status, as low concentration treatment showed higher site-specific phosphorylation revealed by specific phospho-tau (S396) antibody.
Moreover, in this lab, the microtubule binding assay of cell samples was established successfully using tau-352 as a positive control and only MT as a negative control (Figure 3). For both curcumin and LiCl treatment, microtubule-binding activity was inhibited, as demonstrated by the microtubule binding assay. In this experiment, curcumin treatment did not show microtubule binding capabilities but inhibited the binding similar to that in previous studies46,47, whereas it was effective to inhibit site-specific tau phosphorylation at higher concentration. In Figure 1, 10 µM curcumin treatment resulted in minimal expression of phospho-tau compared with control but higher expression than total tau. However, microtubule-binding capacity of colorectal cancer tau after curcumin treatment as well as low concentration of LiCl treatment was decreased in the untreated sample. Site-specific tau phosphorylation effects microtubule binding and self-aggregation48. Tau phosphorylation at proline-rich region impeded microtubule-binding properties whereas C-terminal region increased these properties, and both regions along with MT-binding region lessened its binding properties by about 70% and disordered the microtubules48. Some other factors as well as other site-specific phosphorylation of tau might be involved for this microtubule destabilization of colorectal cancer tau treated with curcumin or LiCl.
A straightforward protocol using small amounts of primary and secondary antibodies enabled localization of tau in colorectal cancer cell lines following curcumin treatment (Figure 4). Results showed that tau had translocated to the nucleus following curcumin treatment, a finding similar to earlier studies reporting that nuclear tau is a key player in neuronal DNA protection in neurodegenerative diseases such as Alzheimer's disease49,50.
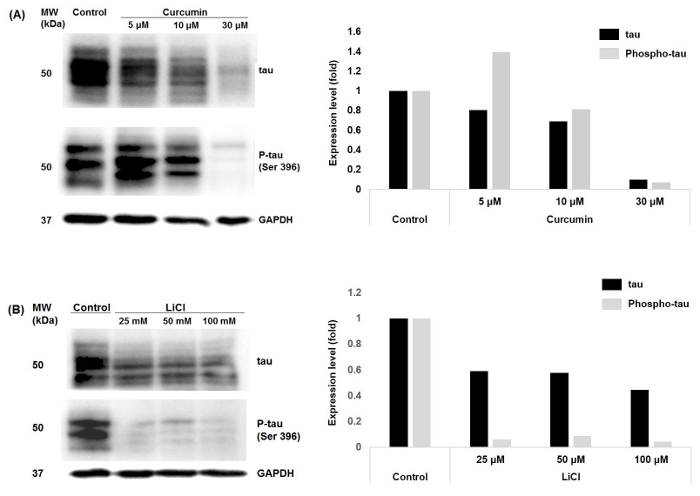
Figure 1: Tau and phospho-tau expression in colorectal cancer cells following curcumin or LiCl treatment. Samples extracted from control cells or cells treated for 24 h with three different concentrations of (A) curcumin and (B) LiCl were resolved on 10% polyacrylamide gels by SDS-PAGE and probed with anti-tau or anti-phospho-tau antibody. Densitometry analyses of the Western blot results are presented on the right panel. Please click here to view a larger version of this figure.
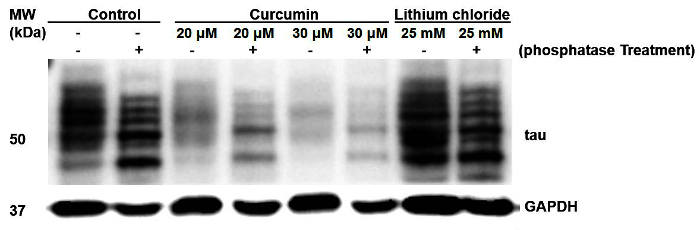
Figure 2: Overall phosphorylation of tau in untreated or differentially treated cell samples detected by the phosphatase assay. Odd lanes show control cell extracts, whereas even lanes show phosphatase treatment of curcumin-treated or LiCl-treated samples. Phosphatase treatment made the tau electrophoretic mobility faster than untreated samples, which contained phosphorylated tau. Please click here to view a larger version of this figure.
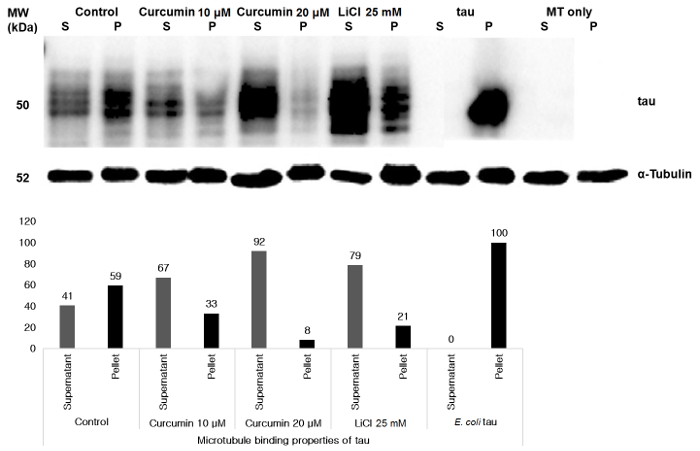
Figure 3: Microtubule binding of tau in colorectal cancer cells detected by the microtubule-binding assay. Control HCT 116 cell samples, curcumin-treated, or LiCl-treated cell samples and E. coli tau were incubated with microtubules as detailed in protocol section 4. Equivalent amounts of supernatant (S) and pellet (P) fractions were immunoblotted with an anti-tau antibody. Negative control lanes containing only MT were run to confirm that MT did not contain any bound endogenous tau. Lower panel shows densitometry analyses of Western blots of individual samples enabling comparison between supernatant (unbound tau) and pellet (bound tau) fractions. Please click here to view a larger version of this figure.
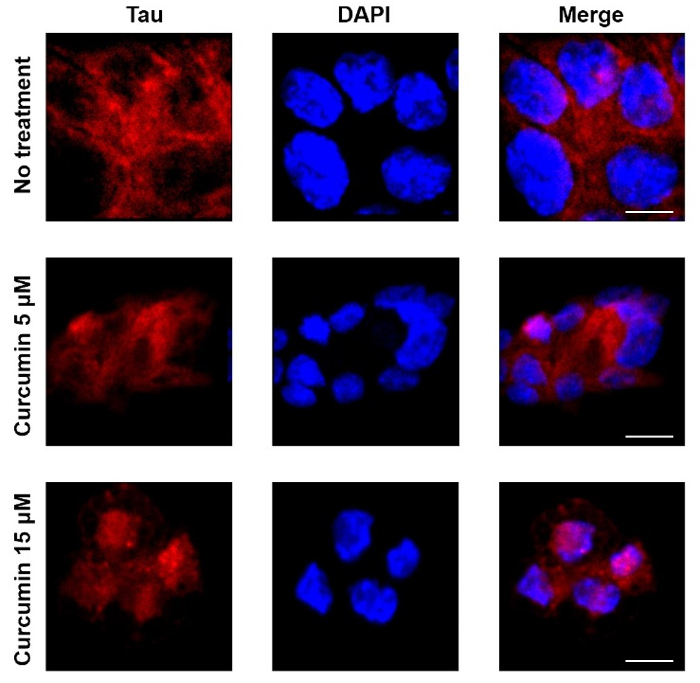
Figure 4: Tau protein localization examined by confocal microscopy.
Tau localized peripherally to the nucleolus in colorectal cancer cells. Left panels show tau localization detected using the anti-tau monoclonal antibody whereas middle panels show nucleus staining by DAPI. Representative examples of treated cells showing tau translocation into the nucleus are also shown. Control cells showed tau mainly around and outside the nuclei, whereas tau in treated cells localized also inside the nuclei. Scale bar = 20 µm. Please click here to view a larger version of this figure.
