Ved at studere arkitekturerne af bærende biologiske strukturer (LBBSs), som shell og knogler, har ingeniører udviklet nye kompositmaterialer, som er både stærk og sej 1. Det har vist sig at de bemærkelsesværdige mekaniske egenskaber af LBBSs og deres bio-inspirerede modparter er relateret til deres indviklede interne arkitekturer 2. Men relationer mellem LBBS arkitekturer og mekaniske egenskaber er ikke fuldt forstået. Måling af en LBBS mekanisk svar er det første skridt mod forståelse hvordan dens arkitektur forbedrer dens mekaniske egenskaber.
Det er imidlertid vigtigt, at typen test bruges til at måle en LBBS mekanisk svar er i overensstemmelse med dens mekaniske funktion. For eksempel da fjer skal understøtte aerodynamiske belastninger, er en fjer rachis primære funktion at give bøjnings stivhed 3. Derfor foretrækkes en bøjning test fremfor et enakset spænding test til at måle dens mekaniske svar. I virkeligheden, mange LBBSs — såsom fjer rachises 3, græs stammer 4og knoglesplinter 5,6,7,8— primært deformeres ved at bøje. Dette skyldes, at disse LBBSs er slank –dvs., deres længde er meget større end deres bredde eller dybde. Imidlertid udføre bøjning test af disse LBBSs udfordrende fordi de styrker og forskydninger, kan modstå før svigtende spænder fra 10-2 til 102 N og 10-4 til 10-3 m, henholdsvis 3 , 4 , 5 , 7 , 8. den enhed, der bruges til at udføre disse mekaniske test bør derfor have kraft og forskydning opløsninger af ≈10-5 N og ≈10-7 m (dvs. 0,1% af sensorens maksimale målelige kraft og forskydning), henholdsvis.
Kommercielt tilgængelige, store skala, mekanisk prøvning systemer typisk ikke kan måle kræfter og forskydninger med denne beslutning. Mens atomic force microscope-baserede 9,10 eller microelectromechanical systemer-baserede 11 test enheder har tilstrækkelig opløsning, den maksimale kraft (respektive forskydning) de kan måle er mindre end den maksimal kraft (respektive forskydning), LBBS kan modstå. Derfor, for at udføre bøjning tests af disse LBBSs, ingeniører og forskere skal stole på specialbyggede mekaniske test enheder 5,7,12,13. Den primære fordel ved disse specialbyggede enheder er, at de kan rumme store intervaller af styrker og forskydninger. Men, opførelse og drift af disse enheder er ikke godt dokumenteret i litteraturen.
En protokol, der er beskrevet for tre-punkts bøjning test bruger specialbyggede mekaniske test udstyr, der kan måle tvinger spænder fra 10-5 til 101 N og forskydninger spænder fra 10-7 til 10-2 m. Tekniske tegninger, herunder alle dimensioner af komponenter af den mekaniske test enhed leveres i det supplerende materiale. Den primære fordel ved denne mekaniske test enhed er, at kraft og forskydning intervaller let kan justeres så de passer til forskellige LBBSs. Enhedens Funktionsprincip er ligner en atomic force microscope 9. I denne enhed, modellen er placeret på tværs af en grøft, skåret i en rustfri plade (Se figur 1A-C). Span af renden er målt fra optiske micrographs skal 1278 ± 3 µm (gennemsnit ± standardafvigelse; n = 10). Skyttegrav kanter støtte modellen under en bøjning test (jf. figur 1 cog D). Denne prøve fase er knyttet til en tre-akse oversættelse fase og anbringes under en aluminium kile, Kilen er placeret midtvejs på tværs af den rende span (Se figur 1C). Ved at flytte fase den 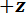 retning (Se figur 1A, og C), modellen er skubbet ind i Kilen forårsager modellen til at bøje.
retning (Se figur 1A, og C), modellen er skubbet ind i Kilen forårsager modellen til at bøje.
Vi henviser til kile som belastning punkt tip (LPT) og del af den enhed, der indeholder kile som belastning (LP). LP er knyttet til slutningen af en cantilever, hvis deplacement er målt af en fiber optic forskydning sensor (FODS). FODS udsender infrarødt lys, hvilket afspejles af et spejl, beliggende på oversiden af LP (Se figur 1B) og modtaget af en optisk fiber i til FODS. En ≈5 mm firkantet stykke plade en poleret silicium anvendes som LP spejl og er fastgjort til LP med epoxy. FODS måler forskydninger ved at sammenligne intensiteten af den udsendte og reflekteret lys. Cantilever stivhed og forskydning er bruges til at beregne kraften, 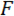 , erfarne af kile på grund af dens interaktion med modellen. Cantilever forskydning er også bruges til at beregne fordrivelse af modellens tværsnit under wedge,
, erfarne af kile på grund af dens interaktion med modellen. Cantilever forskydning er også bruges til at beregne fordrivelse af modellens tværsnit under wedge, 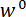 . Cantilever-baserede Kraftmålerne har været brugt i en række mikro – og makro-skala mekaniske test undersøgelser 10,11,12,13,14. Den specifikke design præsenteres her er tilpasset fra en mekanisk prøvningsanordningen anvendes til at udføre selvklæbende kontakt eksperimenter 14. Et lignende design har også været anvendt i et kommercielt tilgængelige mikro-tribometer 15,16.
. Cantilever-baserede Kraftmålerne har været brugt i en række mikro – og makro-skala mekaniske test undersøgelser 10,11,12,13,14. Den specifikke design præsenteres her er tilpasset fra en mekanisk prøvningsanordningen anvendes til at udføre selvklæbende kontakt eksperimenter 14. Et lignende design har også været anvendt i et kommercielt tilgængelige mikro-tribometer 15,16.
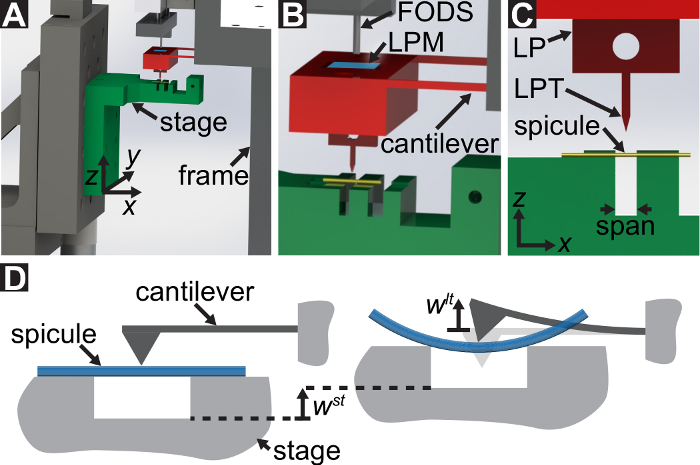
Figur 1: oversigt over den specialbyggede mekaniske prøvningsanordningen. (A) A computer aided design rendering af enheden. Fase komponenter er markeret med grønt. Den kraft sensing halvfabrikata (cantilever, belastning punkt (LP)) er fremhævet med rødt. (B) en forstørret visning af (A). LP spejl er vist med blåt på oversiden af LP under til FODS og er mærket LPM. (C) det koordinatsystem, der bruges til at beskrive bevægelse af oversættelse fase. Ved udjævning the fase i trin 1.9 i protokollen, den retning er lavet til at falde sammen med vektor vinkelret på overfladen af LP spejl. (D) A skematisk af den tre-punkts bøjning konfiguration viser deformation af spicule og de målte forskydninger 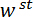 , og
, og 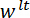 . Venligst klik her for at se en større version af dette tal.
. Venligst klik her for at se en større version af dette tal.
Enhedens funktioner er bevist ved at udføre tre-punkts bøjning test på de skeletale elementer af marine svampen Euplectella aspergillum6,7. Denne svamp skelet er en forsamling af filamenter, kaldet knoglesplinter (Se figur 2A). Knoglesplinter er ≈50 µm tykt og består primært af silikater 6. Biosilica-baserede knoglesplinter findes i svampe tilhører klasserne Demospongiae, Homoscleromorpha og Hexactinellida. Svampe, såsom E. aspergillum, der tilhører klasse Hexactinellida er også kendt som “glas svampe.” Mens knoglesplinter af glas svampe er sammensat primært af silica, har det vist sig at silica ofte indeholder en organisk grundsubstans, sammensat af enten kollagen 17,18 eller kitin 19,20 , 21. denne organiske matrix spiller en vigtig rolle i silica biomineralization 18,20. Desuden i nogle knoglesplinter fungerer den organiske matrix også som en skabelon for biomineralization af calcium 22. Ud over at blive fordelt inden for silica, kan den organiske matrix også danne forskellige lag, at partitionerer den spicule silica i koncentriske, cylindrisk lamellae 6,23. Det har vist sig at denne koncentrisk, gråt arkitektur kan påvirke knoglesplinter deformation adfærd 6,7,8,24,25,26 . Derfor knoglesplinter mekaniske egenskaber bestemmes af en kombination af deres kemi (dvs., den kemiske struktur af silica-protein komposit) og deres arkitektur 27. Både den kemiske struktur og arkitektur af glas svamp knoglesplinter er stadig under efterforskning 24,28,29.
De fleste af knoglesplinter i E. aspergillum er cementerede sammen til at danne en hård skelet bur. I bunden af skelettet er der imidlertid en TOT af meget længe (≈10 cm) knoglesplinter kendt som anker knoglesplinter (Se figur 2A). Vi beskriver protokol til tre-punkts bøjning test på små sektioner af anker knoglesplinter.
I trin 1 i protokollen beskrevne procedure for samling og justering af komponenterne i den specialbyggede mekaniske test enhed. Trin 2 og 4 i denne protokol giver instruktioner til generere kalibreringsdata bruges til at beregne styrker og forskydninger i bøjning test. De skridt til at forberede en del af en spicule og montere det til test Armaturet er beskrevet i trin 3. Procedure for udførelse af bøjning test på afsnittet spicule er beskrevet i trin 5. Endelig, i afsnittet Repræsentant resultater kalibrering oplysninger indhentet i trin 2 og 4 er bruges sammen med bøjning test oplysninger indhentet i trin 5 til at beregne og .

Figur 2: Procedure for skæring og inspicere E. aspergillum knoglesplinter. (A) et skelet af E. aspergillum. Fritstående anker knoglesplinter TOT er vist i bunden af skelettet. Skalalinjen er ~ 25 mm. (B) en enkelt anker spicule er holdt på plads på et objektglas med et #00000 rød Zobel pensel og sectioned ved hjælp af et barberblad. Skalalinjen er ~ 12 mm. (C) en del af en E. aspergillum spicule placeret på tværs af renden på prøve scenen. Skyttegrav kanter og grøft ridge er fremhævet i krikand og orange, henholdsvis. Spicule er skubbet mod renden højderyg til at sikre, at dens akse er vinkelret på renden kanter. (D) A Mikrograf af en spicule, der passerer inspektion fremgangsmåden i trin 3.4 i den protokol, som beskriver, hvordan at afgøre, om et spicule afsnit er beskadiget og skal kasseres. (E) A Mikrograf af en spicule, der indeholder mange revner og mangler store dele af silica lag, der ikke ville kontrolproceduren beskrevet i trin 3.4 i protokollen. Skalere barer = 250 µm (C), 100 µm (D) og 100 µm (E). Venligst klik her for at se en større version af dette tal.
