Adult cardiomyocytes in culture: Figure 1 shows an overview of freshly isolated adult cardiomyocytes 2 h after the last washing. Approximately 75% of all cardiomyocytes had a rod-shaped morphology. The remaining 25% showed an unusual appearance with a round morphology and no detectable intact cell membrane (Figure 1). At the end of cultivation (day 6), up to 15% of all cardiomyocytes showed spreading, about 10% remained in a round morphology without pseudopodia-like structures, and 75% of all cardiomyocytes presented an unusual appearance with an irregular surface and without a detectable intact cell membrane (data not shown).

Figure 1: Overview of freshly isolated rat cardiomyocytes. The fraction of freshly isolated cardiomyocytes which showed a rod-shaped morphology amounted to 75% of cardiomyocytes, on average. The remaining 25% of cells presented an unusual appearance with an irregular surface and no detectable intact cell membrane. Recording was conducted by light microscopy 2 h after washing the freshly isolated cardiomyocytes. Light microscopy 2X magnification. Please click here to view a larger version of this figure.
With light microscopy, freshly isolated ARVC appeared rod shaped and around 100 µm in size (Figure 2A). Freshly isolated ARVC that contract spontaneously were not calcium tolerant. All cells that were round and without a detectable intact cell membrane were damaged and not viable (Figure 2A-B). In the following days, most of the rod shaped ARVC lost this morphology. Cells got rounded with a detectable intact cell membrane. These ARVC were viable. Starting at day three the latter cells formed pseudopodia-like structures. Some of these ARVC kept their rounded appearance during spreading (Figure 2B). Others converted into flat, polymorphic ARVC (Figure 2B).
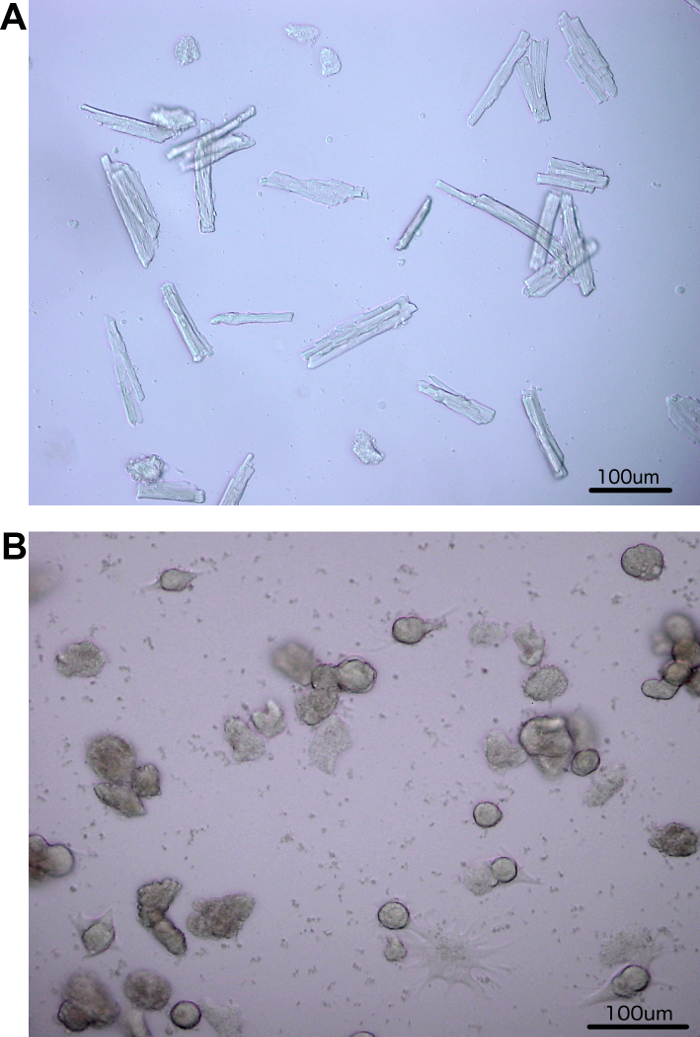
Figure 2: Isolated rat cardiomyocytes. (A) Freshly isolated ARVC were typically rod-shaped. (B) After six days in culture, pseudopodia-like structures (spreading) were clearly detectable in the now rounded ARVC. Some ARVC completely changed to a widespread morphology. ARVC with an unusual appearance displayed an irregular surface and no detectable intact cell membrane. Light microscopy 10X magnification. Please click here to view a larger version of this figure.
Freshly isolated ARVC were typically rod shaped with a clearly visible cross striation (Figure 3, Day 0). Changes in cell morphology were observed during the following days in culture. First, ARVC lost all their contractile elements (Figure 3, Days 1 and 2). This was followed by a reformation, implicating de novo sarcomerogenesis. The reformation was preceded by the formation of pseudopodia-like structures (spreading, Figure 3, Days 3 to 6). De novo sarcomerogenesis started with the appearance of actin stress fibers (Figure 3, Day 3). Additionally, actin bundles appeared in the perinuclear region and formed newly assembled sarcomeres (Figure 3, Days 4 and 5). The latter grew along the preformed actin stress fibers into the periphery (Figure 3, Day 6). At the end of the cultivation period (Day 6a), a typical cross striation from newly assembled sarcomeres in the spread ARVC was observed.
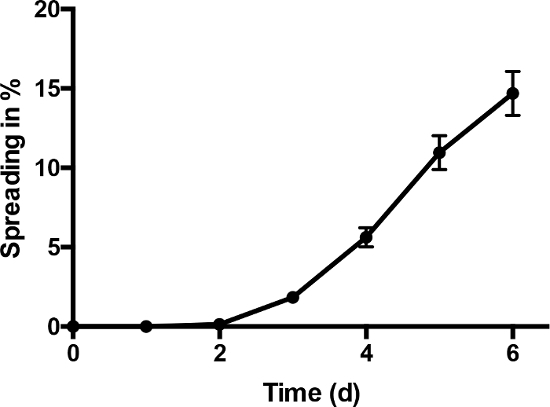
Figure 3: Fluorescence staining. The de- and re-differentiation of ARVC in culture with 20% FCS is shown. Freshly isolated ARVC with their typical rod shape (Day 0) became round by degrading sarcomeres during the first days of culture (Day 1). They lost all their contractile elements (Day 2) followed by formation of pseudopodia-like structures (spreading; Days 3 – 5) and subsequent reformation of their contractile elements indicating de novo sarcomerogenesis (Day 6). At day six in culture, cross striation was clearly detectable again (Day 6a). Staining with Phalloidin-TRITC according to the manufacturer's protocol; "arrows": pseudopodia-like structures (example shown); *: actin bundles in the perinuclear region (exemplary shown). Parts of this figure are published in11. Please click here to view a larger version of this figure.
Figure 4 displays the kinetic of the spreading process during cultivation. The fraction of ARVC showing pseudopodia-like structures at each time of examination is given as spreading in % (Figure 4). Spreading started around day three and increased constantly during the time of cultivation. 14.7% ± 1.39% of all counted ARVC showed pseudopodia-like structures after six days in cultivation.
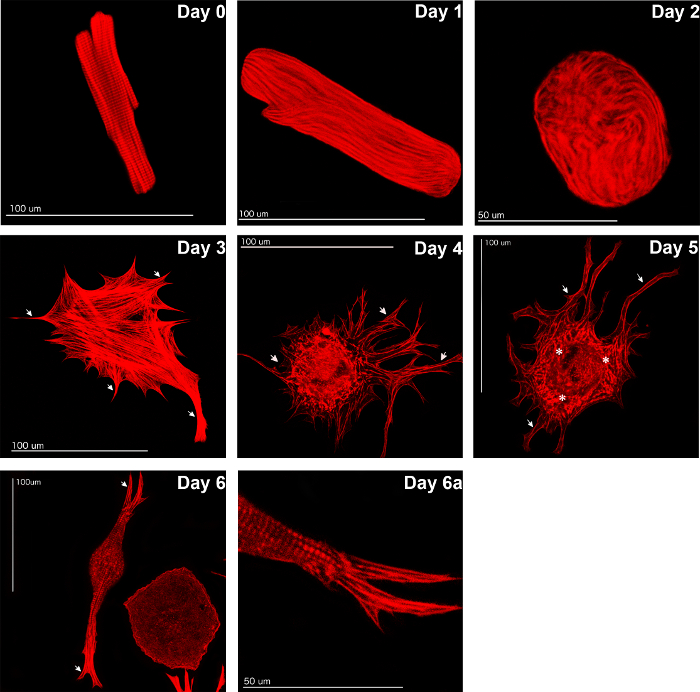
Figure 4: Spreading kinetic increase in cardiomyocytes with pseudopodia-like structures normalized to all counted cardiomyocytes (spreading in %) during six days of cultivation time (n = 33 cell preparations). Data are presented as means ± SEM. This figure is published in11. Please click here to view a larger version of this figure.
Effect of ionomycin on the spreading of ARVC: The isolation and cultivation of ARVC is a calcium sensitive process1,8. Treatment of ARVC with ionomycin (1 µM), which increases intracellular calcium concentration, caused a significant (p ≤0.01) decrease in the formation of pseudopodia-like structures compared to controls (Figure 5). When compared directly, 17.19% ± 2.45% of all counted ARVC showed spreading under control conditions but only 9.87% ± 2.77% of all counted ARVC formed pseudopodia-like structures in the presence of ionomycin (day 6 of cultivation). Thus, ionomycin reduced spreading by 42.58%.
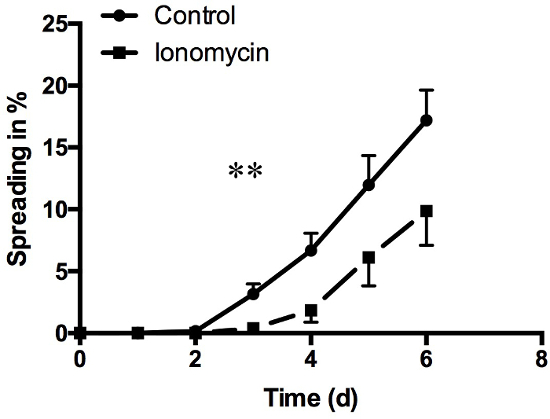
Figure 5: Spreading kinetics under treatment with ionomycin. Treatment with ionomycin (1 µM) at day 0 caused a highly significant reduction in cell spreading compared to control. Data are presented as means ± SEM; n = 4 cell preparations; Mann-Whitney-U test; * p ≤0.05; ** p ≤0.01 Please click here to view a larger version of this figure.
Additionally, ionomycin increased the percentage of ARVC with an unusual appearance compared to control conditions (Figure 6). At day six, 71.11% ± 4.65% of all counted ARVC treated with ionomycin showed an unusual appearance. However, under control conditions, only 51.35% ± 3.55% of the ARVC were categorized in this group.
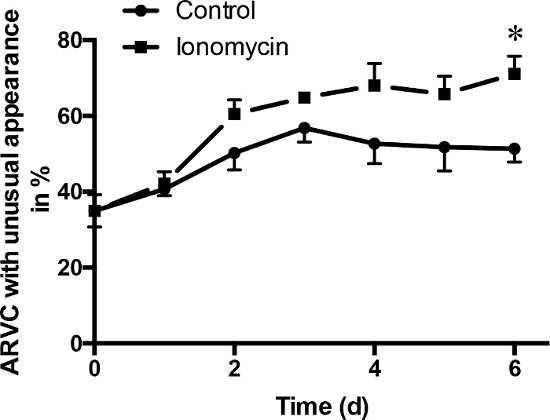
Figure 6: ARVC with an unhealthy appearance. Treatment with ionomycin (1 µM) at day 0 caused a significant increase in the number of ARVC, which showed an unusual appearance. At day 6, the difference between control and ARVC treated with ionomycin was significant. Data are presented as means ± SEM; n = 4 cell preparations; Mann-Whitney-U test; * p ≤0.05 Please click here to view a larger version of this figure.
At day 3 of cultivation, under treatment with ionomycin, qRT-PCR revealed a decrease in mRNA expression of β-MHC (p ≤0.01) and OSM, which both play a distinct role in the de-differentiation of ARVC (Figure 7A and C). Swiprosin-1, a marker for re-differentiation of ARVC was significantly downregulated, too (Figure 7B).
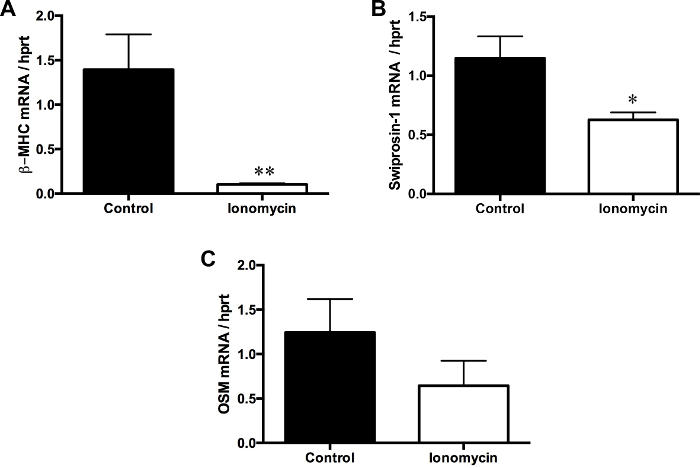
Figure 7: De- and re-differentiation of cultivated ARVC under treatment with ionomycin
(1 µM) at day 0 caused a decreased mRNA expression of oncostatin M (OSM) and β-MHC, which both play key roles in the de-differentiation of adult cardiomyocytes. Additionally, mRNA expression of Swiprosin-1, a key player in the re-differentiation of adult cardiomyocytes, was also decreased by ionomycin treatment. Day 3 of cultivation; Data are presented as means ± SEM; n = 30 cell culture plates per group; Mann-Whitney-U test; * p ≤0.05; ** p ≤0.01 Please click here to view a larger version of this figure.
| Pre-plating medium |
| 20 mL CCT medium |
| 2 % Vol. Penicillin/Streptomycin (400 μL) |
| 4 % Vol. FCS (800 μL) |
| Plating medium |
| 20 mL CCT medium |
| 2 % Vol. Penicillin/Streptomycin (400 μL) |
| Washing medium |
| 20 mL CCT medium |
| 2 % Vol. Penicillin/Streptomycin (400 μL) |
| Note: 4 %Vol. FCS in pre-plating medium can be replaced by 1 Vol.-% laminin (0.5 μg/cm2). Additionally, for cultivating cardiomyocytes for several days add 20 Vol.-% FCS to the washing medium. Store plating medium and washing medium by 4-8 °C until using. |
Table 1: Culture media used for cardiomyocyte isolation
