Exakt gen redigering utgör hörnstenen i stora biomedicinsk framsteg som innebär utvecklingen av nya strategier för att hantera genetiska sjukdomar. I spetsen för genredigering teknik är metoden att förlita sig på användningen av det clustered regularly –jagnterspaced short palindromic repeats (CRISPR) / Cas9-system som identifierades inledningsvis som en del av bakteriell immunitet mot invasionen av viral arvsmassa (ses i referenser1,2). En stor fördel av CRISPR/Cas9 systemet över andra genredigering verktyg, såsom zink finger nukleotider (motvilligt) och transkription aktivator-liknande effektor nukleaser (TALENs) (ses i referens3), är den relativa enkelheten i plasmiden design och byggandet av CRISPR komponenter — en funktion som har driv utbyggnaden av genredigering från några specialiserade laboratorier till en mycket bredare forskarsamhället. Enkelheten av CRISPR/Cas9 programmering och dess kapacitet för multiplexering prismalås har dessutom ytterligare underblåst dess popularitet som en kostnadseffektiv och lätt att använda teknik. Bland de olika metoderna tillgängliga för forskare att leverera sådana genredigering komponenter till celler, fortfarande virala vektorer överlägset mest populära och effektiva systemet.
Lentiviral vektorer (LVs) har dykt upp som fordonet val leverera komponenterna i CRISPR/Cas9-system i vivo för applikationsmöjligheter4,5,6,7. Flera viktiga funktioner gör LVs ett populärt val för denna process inklusive deras förmåga att infektera både dela och icke-dividera celler, låg immunogenicitet och minimal cellulär toxicitet (ses i referens8). Som ett resultat, har LV-medierad genterapi varit anställd i behandlingar av smittsamma sjukdomar, som HIV-1, HBV och HSV-1, samt korrigering av defekter underliggande mänskliga ärftliga sjukdomar som cystisk fibros och neo-vaskulär makuladegeneration 4 , 5 , 7 , 9 , 10 , 11. Dessutom LVs effektivt har ändrats för att utföra multiplex gen redigering på distinkt genomisk lokus använder en enda vector systemet12.
Men den inneboende egenskapen hos LVs att integrera i värd genomet kan vara mutagena och ofta svårigheter deras nytta som transgenens leveransfordon, särskilt i kliniska inställningar. Dessutom eftersom stabilt-integrerade LVs express deras transgener på hållbart höga nivåer, är detta system dåligt lämpade för leverans av genredigering komponenter såsom CRISPR/Cas9; överuttryck av Cas9-guide RNA (gärna), och liknande proteiner som motvilligt, är associerade med förhöjda nivåer av off-effekter, som inkluderar oönskade mutationer13,14,15,16 , 17 och kan potentiellt öka cytotoxiciteten18. För att uppnå exakt genredigering med minimal off-target effekter, det är därför absolut nödvändigt att konstruera system som möjliggör övergående uttrycket av genen redigering komponenter.
Under de senaste åren har en mängd olika leveransplattformar utvecklats för att övergående express CRISPR/Cas9 i celler16,19,20,21 (ses i referens22). Dessa inkluderar metoder som förlitar sig på direkt införa renat Cas9 tillsammans med lämplig guide RNASEN i celler, vilket visade sig vara mer effektiva på riktade genredigering jämfört med plasmidmedierad transfection16. Studier har visat att ribonukleoprotein (RNP) komplex bestående av guide RNA/Cas9 partiklar vänds snabbt efter medla DNA klyvning på sina mål, vilket tyder på att kortsiktiga uttryck av dessa komponenter är tillräckligt för att uppnå robust genen redigering16. Möjligen, icke-integrering virala vektorn plattformar såsom adeno-associerade virala vektorer (AAVs) kan ge ett lönsamt alternativ för att leverera genredigering maskiner till celler. Tyvärr AAV förhoppningsvis ha betydligt lägre förpackning kapacitet än LVs (< 5kb), som allvarligt hämmar deras förmåga att paketera flera komponenter CRISPR toolkit inom en enda vektor (ses i referens8). Det är värt att notera att tillägg av föreningar som hämmar Histon deacetylases (e.g., natrium butyrate23) eller hindra cellcykeln (t.ex., koffein24) har visat sig öka lentiviral titrar. Trots de senaste framsteg hindras de övergående uttryck system utvecklat hittills fortfarande av flera brister, såsom lägre produktionseffektivitet, vilket leder till minskad viral titrar och låg transduktion effektiviteten av de virus som genereras genom sådana strategier25.
Integras-brist lentiviral vektorer (IDLVs) utgör ett stort framsteg i utvecklingen av gen-leverans fordon, eftersom de kombinerar LVs förpackning förmåga med fördelen av AAV-liknande episomal underhåll i celler. Dessa funktioner hjälpa IDLVs till stor del kringgå de viktiga frågor i samband med att integrera vektorer, vis-à-vis kontinuerlig överuttryck av potentiellt genotoxiska element och integration-medierad mutagenicitet. Det visades tidigare att IDLVs framgångsrikt kan ändras för att förbättra episomal gen uttryck26,27. När det gäller IDLV-medierad CRISPR/Cas9 leverans begränsar låg produktion titrar och lägre uttryck för episome-burna genomen i förhållande till integras-kunnig lentiviral system deras användbarhet som bona fide verktyg för att leverera editering transgena konstruktioner. Vi har nyligen visat att både transgenens uttryck och viral titrarna är associerade med IDLV produktionen har förbättrats avsevärt genom införandet av bindande platser för Transkriptionfaktorn Sp1 i viral uttryck kassett28. De modifierade IDLVs stöds kraftfullt CRISPR-medierad gen redigering både in vitro- (i HEK-293T celler) och i vivo (i efter mitotiska hjärnans nervceller), medan inducerande minimal off-target mutationer jämfört med motsvarande ICLV-medierad system28. Sammantaget utvecklade vi en roman, kompakt, allt-i-ett CRISPR toolkit transporteras på en IDLV plattform och beskrivs de olika fördelarna med att använda sådant leveransfordon för förbättrade genen redigering.
Här är protokollet produktion av IDLV-CRISPR/Cas9 systemet beskrivs, inklusive de olika steg som ingår i församlingen, rening, koncentration, och titrering av IDLVs, samt strategier för att validera genredigering effekten av dessa vektorer. Detta protokoll är enkelt skalbart för att möta behoven hos olika undersökare och är utformad att framgångsrikt generera LV vektorer med titrar i spänna av 1 x 1010 transducing enheter (TU) / mL. De vektorer som genereras genom detta protokoll kan utnyttjas för att effektivt angripa flera olika celltyper, däribland svårt-att-transduce embryonala stamceller, hematopoetiska celler (T-celler och makrofager) och odlade och in vivo– injicerade nervceller. Protokollet är dessutom lika väl lämpad för produktion av integras-behöriga lentiviral vektorer i liknande kvantiteter.
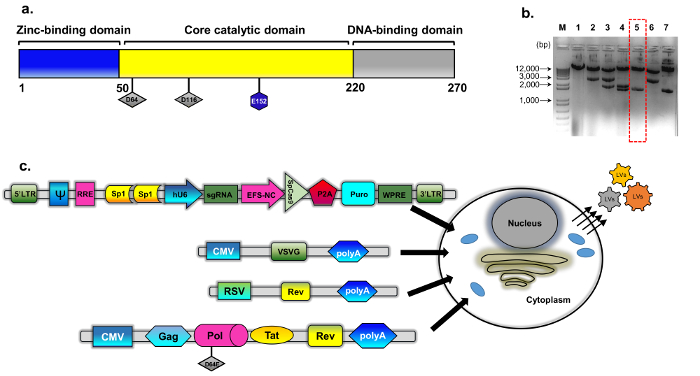
Figur 1: IDLV förpackningar. (a) Schematisk bild av vildtyp integras protein (b) modifierad plasmiden härleddes från psPAX2 (se metoder, plasmid konstruktion för detaljer). Representant agaros gel bild av kloner screenas för muterade integras kloner. DNA-prover som tillagas med ett mini-kit med standard plasmid-DNA med isolering analyserades av matsmältningen med EcoRV och SphI. Korrekt rötas klonen (nummer 5, streckad röd ruta) bekräftades ytterligare genom direkt (Sanger) sekvensering för D64E substitution i INT. Integras-bristfällig förpackning kassetten hette pBK43. (c) Schematisk av protokollet övergående transfection anställd för att generera IDLV-CRISPR/Cas9 vektorer, visar 293T celler transfekterade med VSV-G, förpackningar och transgenens kassetter (Sp1-CRISPR/Cas9 allt-i-ett plasmid). Viruspartiklar som bud ut från cellmembranet innehåller fullängds RNA av vektor (uttryckt från transgenens kassett). Den andra generationen av systemets IDLV-förpackning användes, vilket inkluderar de reglerande proteinerna Tat och Rev. Rev uttryck kompletteras vidare från en separat kassett (RSV-REV-plasmid). ABBREV: LTR-lång-terminal upprepa, VSV-G, vesikulär stomatit virus G-protein, pCMV-cytomegalovirus arrangören; Rous sarkom virus (RSV) arrangören; RRE-(Rev svar Element). Andra reglerande element på uttrycket kassetten omfattar Sp1-bindande platser, Rev Response element (RRE), murmeldjur hepatit Virus från föreskrivande Element (WPRE), en core-töjning faktor 1α promotor (EFS-NC), vektor förpackningen elementet ψ (psi), human Cytomegalovirus (hCMV) arrangören och mänskliga U6 arrangören. Klicka här för att se en större version av denna siffra.
