Vi beskriver en samlet intern refleksion fluorescens-antal og lysstyrke (TIRF-N & B) Imaging tilgang til bestemmelse af den gennemsnitlige oligomeriske tilstand af receptor molekyler i plasma membranen af levende celler, sigter mod at forbinde receptoren forsamling dynamik til proteinerne biologiske funktion (figur 1).
Ved ekstracellulær ligand binding initierer receptorer det intracellulære signal transduktion afhængigt af deres kropsbygning, oligomerisering, potentielle Co-receptorer og membran sammensætning. På trods af betydningen og allesteds nærende receptor oligomerisering, anerkendt som en vigtig begivenhed i cellulære signalering1,2,3,4,5,6, 7, få metoder kan detektere klyngedannelse begivenheder og måle graden af klyngedannelse eksperimentelt8,9. Den confokale volumen (x, y ≈ 300 Nm, z ≈ 900 nm) er utilstrækkeligt løst til at bevise molekylære interaktion og støkiometri, selv efter optimering af billedet restaurering algoritmer10. Under enheds sammensætningen af protein oligomerer kan ikke løses på et rent geografisk grundlag, selv ved super opløsnings metoder ved x, y-opløsning på 20-70 nm såsom PALM11, storm12og sted13. Desuden kan deres tidsmæssige opløsning (i rækkefølgen af minutter pr. billede) ikke følge kinetikken i intervallet fra sekunder. Enkelt molekyle Step-blegning løser kun støkiometri af protein oligomerer, hvis de er immobile14.
En af de mest alsidige metoder til at måle tæthed og oligomerisering af fluorescently mærkede proteiner inden for enkeltbilleder er den rumlige intensitets fordelings analyse (SpIDA), som er afhængig af rumlig prøvetagning. Det gælder både kemisk faste og levende celler, og gør det muligt at analysere flere områder af interesse for cellen samtidig ved hjælp af standard Fluorescens mikroskopi15. Alternativt, moment metoder, såsom fluorescens-korrelations spektroskopi (FCS)16, photon Counting histogram (PCH)17, og tal og lysstyrke (N & B)18,19, er egnet til kvantitativ oligomer Målinger. Disse metoder analyserer fluorescens intensitets udsving, der kan observeres i tide, når fluorophorerne diffus ind og ud af en belysnings volumen. Amplituder af intensitets svingningerne kan entydigt beskrives ved den molekylære lysstyrke af fluoroforet (ε) og det gennemsnitlige antal fluorophorer (n) i belysnings volumenet17 (figur 2). Typisk kan diffusions koefficienten for fluorophorerne og det gennemsnitlige antal molekyler (omvendt relateret til G (0)-værdien) inden for belysnings volumenet opnås af FCS20. Men da diffusions tiden kun skalerer med den kubiske rod af massen, er FCS ikke tilstrækkelig følsom til at påvise ændringer i molekylmasse21. I praksis kan en enkelt farve FCS ikke detektere dimerisering af membranreceptorer. PCH løser blandinger af forskellige oligomerer nøjagtigt. Ved hjælp af mere end to øjeblikke af amplitude fordelingen, det registrerer molekyler af forskellig lysstyrke, der indtager den samme belysning volumen. Scanning FCS22 og udviklinger, såsom den interessante par-korrelation af Molekylær lysstyrke (pcomb) tilgang23, indført for at udvide anvendelsesområdet for fluorescens korrelations metoder i biologiske systemer24 , forblive enkelt punkt metoder, der mangler evnen til hurtige målinger i et stort område af en celle, kræver mange på hinanden følgende observationer på hver pixel og data erhvervelse i rækkefølgen af sekunder.
N & B er en forenklet version af PCH, der kun betragter det første og andet øjeblik af amplituden i fluorescens fordelingen, nemlig den gennemsnitlige intensitet, , og variansen, σ2 (figur 2)18,19 og på grund af dette, kan den ikke bestemme den molære fraktion af ukendte oligomerer i en blanding, men anslår kun den gennemsnitlige oligomeriserings tilstand af blandingen. Ikke desto mindre har N & B den fordel at arbejde med relativt mindre tidsserier af billeder af levende celler end PCH på en pixel-for-pixel basis, blot ved at overvåge udsving på tidspunktet for fluorescens intensitet. Da N & B reducerer tiden pr. pixel til nogle få mikrosekunder, kan den følge en hurtig oligomeriserings kinetik over store celleområder, hvilket giver mulighed for at erhverve billeder på en tidsskala i sekunder i mikroskopi af raster scanning (f. eks. confocal, 2-photon) og millisekunder i Kamerabaseret mikroskopi (f. eks. TIRFM).
Flere rapporter har vist, at N & B er i stand til at kvantificere antallet af under enheder i protein klynger ved at afbilde udvidede celleområder. Der blev fundet paxillin-EGFP-klynger på vedhæftnings stederne i CHO-K1-cellerne25, og den intracellulære sammenlægning af det patogene Httex1p peptid blev beskrevet i cos-7-cellerne26. N & B blev ansøgt om at følge den ligand-drevne oligomerisering af ErbB-receptoren27og virkningen af ligand-FGF21 på Klothob (klb) og FGFR1c i Hela-cellerne28. Kombinationen af TIRF Imaging og N & B analyse blev brugt til at vise, at dynamin-2 primært er tetrameric i hele cellemembranen29. Vi anvendte N & B til både raster scanning og tirf billeder til at bevise ligand-drevet denne af uPAR og FGFR1 celle membranreceptorer30,31.
Fluorescens korrelations metoder, såsom N & B, FCS og PCH, er baseret på forestillingen om, at antallet af partikler i et åbent volumen følger en Poisson-fordeling. Da kun de fotoner, som fluorophorerne udsender, kan påvises, er middelværdien for en målt fluorescens intensitet versus tid i en pixel i billedet  , produktet af det gennemsnitlige antal fluorophorer i belysnings volumen n og deres Molekylær lysstyrke, ε17:
, produktet af det gennemsnitlige antal fluorophorer i belysnings volumen n og deres Molekylær lysstyrke, ε17:

hvor ε udtrykkes som antallet af fotoner, der udsendes pr. tidsenhed (konventionelt pr. sekund) pr. molekyle, når molekylet er i midten af belysnings volumenet.
Lysstyrke er en egenskab for hver fluoroforet i en given erhvervelse oprettet, mens intensitet er summen af alle bidrag fra alle fluorophorer. I biologiske konkurrencer, vil lysstyrken stige med stigningen i antallet af fluorophorer, der svinger sammen, giver oplysninger om oligomerisering tilstand af fluorescently-mærkede protein. Udsvingene amplituder ved en given pixel måles fra variansen af fluorescens signalet, σ2:

Hvor middelværdien af kvadratet af intensitet  ,, og kvadratet af middelværdien af
,, og kvadratet af middelværdien af  intensitet, er beregnet ud fra de individuelle intensitetsværdier i hver pixel i hver ramme:
intensitet, er beregnet ud fra de individuelle intensitetsværdier i hver pixel i hver ramme:
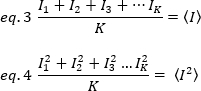
hvor K er antallet af samlede rammer i tidsserierne. Eksperimentelt er det nødvendigt at beregne variansen for hele billedserien, der beskriver spredningen af de individuelle intensitetsværdier ved hver pixel i et enkelt billede omkring middelintensitets værdien. Variansen omfatter alle udsving af forskellig oprindelse. Ved første tilnærmelse kan variansen på grund af sprede-partiklerne i belysnings volumenet, σ20, adskilles fra variansen på grund af detektor støjen, σ2d. De to afvigelser er uafhængige; således, den samlede varians er givet ved deres sum:

Variansen på grund af molekylære udsving i og ud af detekterings volumenet er lineært afhængigt af den molekylære lysstyrke og intensitet:

Omarrangering af EQ. 6 ifølge EQ. 1:

Ifølge den typiske koncept i fluorescens korrelations spektroskopi, ligning 7 hedder det, at variansen på grund af antallet af udsving afhænger af kvadratet af partikel lysstyrken.
Variansen på grund af detektor udsving er derefter en lineær funktion af den detekterede intensitet under den antagelse, at detektoren drives under dens mætningsgrænse19:

I tilfælde af foton tælle detektorer a= 1 og c= 0, således detektor variansen er lig med den gennemsnitlige intensitet:

For at anvende disse begreber til reelle målinger i levende celler, Gratton og kolleger18 definere den tilsyneladende lysstyrke, B, for hver pixel som forholdet mellem variansen over den gennemsnitlige intensitet:

B er den parameter, der måles eksperimentelt. I dette arbejde, tidsserie billeder af FGFR1 receptorer på plasma membranen af HeLa celler er fanget af TIRF mikroskopi og den gennemsnitlige tilsyneladende lysstyrke, B, bestemmes af N & B analyse. Derefter, efter tilsætning af FGF2, sammenhængende tidsserier er fanget for at følge ændringerne i den selv-samling af receptor molekyler i membranen overflade efter stimulation af receptoren med den kanoniske ligand.
Men da detektoren af TIRF mikroskop er et EMCCD-kamera, skal udtrykket for den tilsyneladende lysstyrke ændres som19:

hvor offset er den intensitets forskydning af detekterings elektronikken, der er karakteristisk for detektor indstillingerne. Variansen og den gennemsnitlige intensitet for en analog detektor angives henholdsvis ved:
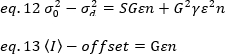
hvor G er analog gevinst i digitale niveauer (DL/photons), S, de digitale niveauer per photon19, er givet ved hældningen af en intensitet versus varians plot for en lyskilde med konstant intensitet (ingen tidsmæssige udsving). Γ-faktoren er relateret til form af pixel detekterings volumen. Ifølge Hassler et al.32er γ faktoren lig med 0,3 for tirf-billeddannelse, der arbejder ved den maksimale gevinst ved detekterings kameraet19. Parametrene offset, S og G er egenskaber for kameraet og mikroskopet. Den tilsyneladende lysstyrke, B, opnås ved at omarrangere EQ. 11 i henhold til EQ. 12 og 13:

Eksperimentelt er ε en kompleks funktion af laser intensitet og systemets detekterings effektivitet. Da B/S ikke desto mindre er lineært afhængig af ε, er det kun vigtigt at bestemme den relative værdi af ε for en given detekterings tilstand:

hvor ε ‘ er proportional med ε. Alligevel udføres en kalibrering ved hjælp af en intern reference.
