I de senere år har betydningen av encellede eksperimenter blitt tydelig. Data fra enkeltceller tillater undersøkelse av celle-til-celle-variasjon, oppløsningen av intracellulære parameterkorrelasjoner og påvisning av cellulær kinetikk som forblir skjult i ensemblemålinger1,2,3. For å undersøke cellulær kinetikk av tusenvis av enkeltceller parallelt, er det nødvendig med nye tilnærminger som gjør det mulig å overvåke cellene under standardiserte forhold over en tidsperiode på flere timer opptil flere dager etterfulgt av en kvantitativ dataanalyse 4. Her presenterer vi Live-cell Imaging of Single-Cell Arrays (LISCA), som kombinerer bruken av mikrostrukturerte matriser med tidsforløpmikroskopi og automatisert bildeanalyse.
Flere metoder for å generere mikrostrukturerte encellede matriser er etablert og publisert i litteratur5,6. Her beskriver vi kort mikroskala plasmainitiert proteinmønster (μPIPP). En detaljert protokoll for enkeltcellet matriseproduksjon ved hjelp av μPIPP finnes også i referanse7. Bruken av encellede matriser gjør det mulig å justere tusenvis av celler på standardiserte adhesjonspunkter som presenterer definerte mikromiljø for hver celle og dermed reduserer en kilde til eksperimentell variasjon (Figur 1A). Encellede matriser brukes til å overvåke tidsforløpene til fluorescerende markører med formål å indikere en rekke cellulære prosesser. Langsiktig mikroskopi i skannetidsforløpmodus gjør det mulig å overvåke et stort område av encellede matriser og dermed prøvetaking av encellede data i høy gjennomstrømning over en observasjonstid på flere timer eller til og med dager. Dette genererer tidslinjestakker med bilder fra hver plassering av matrisen (Figur 1B). For å redusere den store mengden bildedata og trekke ut de ønskede enkeltcellede fluorescenstidskursene på en effektiv måte, kreves automatisert bildebehandling som utnytter plasseringen av celler (Figur 1C).
Liscas utfordring er å tilpasse eksperimentelle protokoller og beregningsverktøy for å danne en analyse med høy gjennomstrømning som genererer kvantitative og reproduserbare data om cellulær kinetikk. I denne artikkelen gir vi en trinnvis beskrivelse av de enkelte metodene og hvordan de kombineres i en LISCA-analyse. Som et eksempel diskuterer vi tidsforløpet for forbedret grønt fluorescerende proteinuttrykk (eGFP) etter kunstig mRNA-levering. eGFP-uttrykk etter mRNA-levering er beskrevet ved reaksjonshastighetsligninger modelleringsoversettelse og nedbrytning av mRNA. Montering av modellfunksjonen for tidsforløpet av eGFP-konsentrasjon til LISCA-avlesningen av fluorescensintensiteten for hver enkelt celle over tid gir de beste estimatene av modellparametere som mRNA-nedbrytningshastigheten. Som et representativt resultat diskuterer vi mRNA-leveringseffektiviteten til to forskjellige lipidbaserte transfeksjonsmidler og hvordan deres parameterfordelinger er forskjellige.
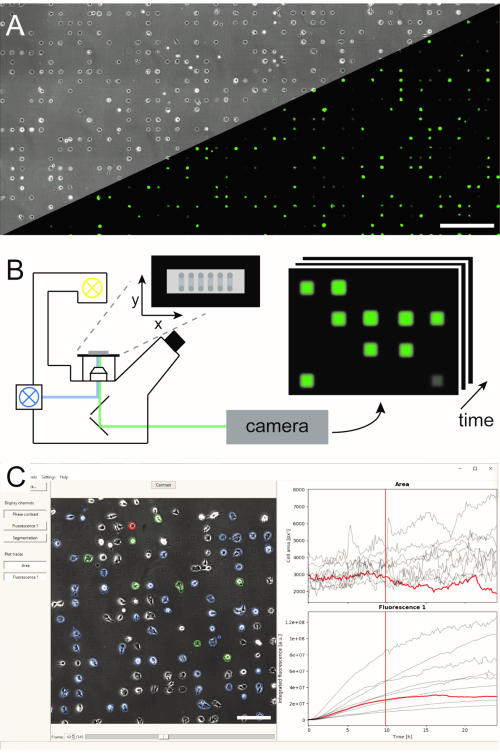
Figur 1: Representasjon av LISCA-arbeidsflyten som kombinerer (A) mikromønstrede encellede matriser (B) som skanner tidsforløpmikroskopi og (C) automatisert bildeanalyse av innspilte bildeserier. Encellede matriser består av et todimensjonalt mønster av celleklebende firkanter med et celleavvisende interspace som fører til et arrangement av cellene på mikropatternen, som det fremgår av fasekontrastbildet samt fluorescensbildet av eGFP som uttrykker celler (A). Hele det mikrostrukturerte området er avbildet i en tidsforløpmodus for skanning gjentatte ganger ved å ta bilder i en sekvens av posisjoner (B). Innspilte bildeserier behandles for å lese ut fluorescensintensiteten per celle over tid (C). Skalastenger: 500 μm (A), 200 μm (C). Klikk her for å se en større versjon av denne figuren.
