Restriction endonucleases (REases) are enzymes that effect sequence-specific double-strand breaks in DNA. The discovery of REases in the 1970s led to the development of recombinant DNA technology, and these enzymes are now indispensable laboratory tools for genetic modification and manipulation1. Type II REases are the most widely used enzymes in this class as they cleave DNA at a fixed location either within or near their recognition sequence. However, there is a great deal of variation among the Type II REases, and they are divided into several subtypes based on particular enzymatic properties rather than being classified according to their evolutionary relationships. Among each subtype, there are frequent exceptions to the classification scheme, and many enzymes belong to multiple subtypes2. Thousands of Type II REases have been identified, and hundreds of them are commercially available.
However, in spite of the diversity among the Type II REases, very few REases have been studied in detail. According to REBASE, the restriction enzyme database established by Sir Richard Roberts in 19753, published kinetics data are available for fewer than 20 of these enzymes. Furthermore, while some REases have been directly observed at the single-molecule level while diffusing along the DNA prior to encountering and binding to their recognition sequence4,5,6,7, there are very few single-molecule studies of their cleavage reaction kinetics. The existing studies either do not report adequate statistics to undertake detailed analysis of the variation in the times at which single cleavage events take place8,9,10 or are not capable of capturing the full distribution of cleavage times11. This type of analysis can reveal the presence of relatively long-lived kinetic intermediates and could lead to better understanding of the mechanisms of REase-mediated DNA cleavage.
At the single-molecule level, biochemical processes are stochastic-the waiting time for a single instance of the process to occur, τ, is variable. However, many measurements of τ can be expected to obey a probability distribution, p(τ), that is indicative of the type of process taking place. For instance, a single-step process, such as the release of a product molecule from an enzyme, will obey Poisson statistics, and p(τ) will take the form of a negative exponential distribution:
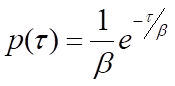
where β is the mean waiting time. Note that the rate of the process, k, will be equal to 1/β, the inverse of the mean waiting time. For processes that require more than one step, p(τ) will be the convolution of the single-exponential distributions for each of the individual steps. A general solution for the convolution of N single-exponential decay functions with identical mean waiting times, β, is the gamma probability distribution:
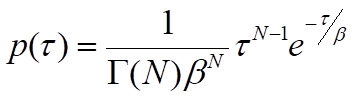
where Γ(N) is the gamma function, which describes the interpolation of the factorial of N-1 to non-integer values of N. Although this general solution can be used as an approximation when the mean waiting times of individual steps are similar, it must be understood that the presence of relatively fast steps will be masked by steps with significantly longer waiting times. In other words, the value of N represents a lower limit on the number of steps12. With an adequate number of waiting-time measurements, the parameters β and N can be estimated by binning the events and fitting the gamma distribution to the resulting histogram or by using a maximum-likelihood estimation approach. This type of analysis can therefore reveal the presence of kinetic steps that cannot be easily resolved in ensemble assays and requires a large number of observations to estimate parameters accurately12,13.
This paper describes a method to use quantum-dot-labeled DNA and total internal reflection fluorescence (TIRF) microscopy to observe hundreds of individual REase-mediated DNA cleavage events in parallel. The design of the assay makes it possible to pool the results of several experiments and can create dwell-time distributions containing thousands of events. The high photostability and brightness of quantum dots permit a 10 Hz time resolution without sacrificing the ability to observe cleavage events occurring even many minutes after the start of the experiment. Good temporal resolution and a broad dynamic range, combined with the ability to collect a large data set, allow accurate characterization of the dwell-time distributions to uncover the presence of multiple kinetic steps in the cleavage pathways of REases, which have turnover rates in the 1 min-1 range. In the case of EcoRV, three kinetic steps can be resolved, all of which have been identified through other means, confirming that the assay is sensitive to the presence of such steps.
Duplex DNA substrates containing the recognition sequence of interest are produced by annealing a biotinylated oligonucleotide to a complementary strand labeled with a single, covalently attached semiconductor nanocrystal quantum dot. These substrates are introduced into a flow channel built on top of a glass coverslip with a lawn of high molecular weight polyethylene glycol (PEG) molecules covalently attached to its surface. The DNA substrates are captured via a biotin-streptavidin-biotin linkage by a fraction of the PEG molecules that have a biotin at their free end. In TIRF microscopy, an evanescent wave that decays exponentially with distance from the glass-liquid interface provides illumination; the penetration depth is on the order of the wavelength of the light used. Under these conditions, only quantum dots that are tethered to the surface by a DNA molecule that has been captured on the functionalized glass surface will be excited. Quantum dots that are free in solution will not be constrained within the illuminated region and therefore will not luminesce. If the DNA tethering a quantum dot to the surface is cleaved, that quantum dot will be free to diffuse away from the surface, and it will disappear from the fluorescence image.
Although many Type II REases are known to bind DNA in the absence of magnesium14, all require magnesium to mediate DNA cleavage15. These REases can bind the surface-immobilized DNA in the absence of magnesium. When magnesium-containing buffer is flowed through a channel with REase prebound to the DNA, cleavage begins immediately, as indicated by the disappearance of quantum dots. The synchronization achieved by prebinding the REase molecules, and then initiating DNA cleavage by introducing magnesium, facilitates measurement of the lag time to the completion of DNA cleavage independently for each molecule in the population of enzymes observed in an experiment. Fluorescein is included as a tracer dye in the magnesium-containing buffer to indicate the arrival of magnesium into the field of view. As no enzyme is included in the magnesium-containing buffer, the lag time from the arrival of magnesium-containing buffer to the disappearance of each quantum dot indicates the time it takes for an REase that is already bound to the DNA to cleave the DNA and release the quantum dot from the glass surface. Quantum dot disappearance happens quickly and results in a sharp decrease in the intensity trajectory, providing a clear indication of the time at which a given DNA molecule is cleaved. The determination of event times is accomplished by mathematical analysis of intensity trajectories, and a typical experiment results in hundreds of identifiable cleavage events. The results of multiple experiments can be pooled to provide adequate statistics to allow estimation of the parameters, N and β, by either nonlinear least-squares or maximum-likelihood analysis.
