Förmågan att modifiera arvsmassan hos myggvektorer av sjukdomar på ett tillförlitligt och reproducerbart sätt har stärkt in vivo funktionell validering av gener och öppnat dörrarna till realiserbara genetiska vektorkontrollstrategier, till exempel de som riktar sig mot Anopheles myggor som överför malaria1.
Tidig mygggenomredigering förlitade sig enbart på transponerbar element (TE)-medierad omvandling, där piggyBac var den vanligaste transposonen i Anopheles2,3,4. Te-integrationens slumpmässiga karaktär kan dock leda till oönskade modifieringar såsom gen knockouts (insertional mutagenesis) och betydande positionseffekter på transgeneuttryck5,6,7,8. Flera infogningar är också vanliga när du använder piggyBac5,9, vilket gör valideringen och isoleringen av transgena linjer med enstaka infogningar mödosamma. Andra nackdelar inkluderar deras potentiella remobilisering, som observerats i bakterien av Anopheles stephensi när de tillhandahåller en källa till piggyBac transposase10,11,12, och deras begränsade storlek på DNA-last (10-15 kb i längd) med omvandling effektivitet minskar med ökande storlek på givaren plasmid13,14.
Webbplatsstyrda integrationsmetoder infördes för att kringgå dessa frågor. Den vanligaste platsstyrda genommodifieringen hos myggor är den som förmedlas av φC31-systemet (figur 1a). Detta drivs av en viral integras som katalyserar rekombinationen mellan två heterospecific anknytning (att) platser som förekommer naturligt i genomet av bakteriofag φC31 (attP) och i Streptomyces bacterium värd (attB)15. Rekombinationen av de två platserna är enkelriktad och resulterar i bildandet av hybridplatser (attL och attR). Rekombinationen av sådana hybridplatser (som leder till DNA-excision) skulle kräva inte bara närvaron av ett aktivt viralt integras utan också en annan fagerkodad rekombinationsfaktor16,17. En stabil integrationsplats skapas därmed som avlastar frågan om potentiell oönskad remobilisering15. Dessutom gör systemet det möjligt att integrera stora laster (t.ex. integrering av >100 kb-konstruktioner rapporterades i D. melanogaster18), vilket avsevärt ökar bärkapaciteten. Integration sker i en enda fördefinierad genomisk locus som kraftigt förenklar valideringen av insättning och parningssystemet för att erhålla en stabil transgen linje. Slutligen tillåter den platsstyrda karaktären av integrationen normalisering av uttryck eftersom alternativa transgener ligger i samma locus och regleras därför inom samma närliggande genomiska sammanhang. En av de viktigaste tillämpningarna av tekniken är den direkta jämförelsen av fenotyper som ges av olika transgener efter införandet i en identisk locus.
Att uppnå φC31-medierad integration innebär två faser: fas I är skapandet av transgena dockningslinjer som transporterar attP-anläggningar, och fas II är den platsstyrda integrationen av en attB-flankerad last i dockningslinjen19. Skapandet av fas I-dockningslinjer har förlitat sig på te-medierad slumpmässig integration av attP-märkta konstruktioner och därmed involverat en inledande mödosam process (inklusive södra blot och omvända PCR-analyser på en-kvinnliga avkomma) för att isolera och validera transgena linjer som bär en enda integrationshändelse i unika, transkriptionellt aktiva och fitnessneutrala genomiska platser. Ändå har flera dockningslinjer för φC31-medierad enkel integration utvecklats och validerats i An. gambiae19,20,21,22 och i An. stephensi23,24,25 (tabell 1). Var och en av dessa linjer varierar när det gäller dockningsplatsens genomiska läge och den stamspecifika genetiska bakgrunden och från dem kan en stor variation av nya transgena linjer skapas. Den komplexa valideringen av TE-medierade integrationer för tillverkning av dockningslinjer kan nu kringgås med CRISPR/Cas9-tekniken26. Detta förlitar sig dock på a priori kunskap om neutral lokus att vara mål och deras omgivande sekvenser.
φC31-medierad integration har tillämpats i stor utsträckning på insektsgenomredigering från modellorganismen D. melanogaster27, till myggorna Aedes aegypti13,28, Ae. albopictus29, An. gambiae19 och An. stephensi24, liksom andra insekter inklusive Ceratitis capitata30 och Bombyx mori31.
En begränsning av φC31-medierad integration, särskilt med tanke på potentiella fältutsläpp för vektorkontroll, är integrationen i mygggenomet hos hela attB-bärande donatorplasmid, inklusive oönskade sekvenser som genmarkörer för antibiotikaresistens och plasmid stamnätkomponenter av bakteriellt ursprung. För att ta itu med detta genomfördes en ändring av standardsystemet, rekombinerasmedierat kassettutbyte (RMCE), som gör det möjligt att exakt ersätta en tidigare integrerad transgen kassett med ett nytt donator-DNA (figur 1b). Detta uppnås genom att använda två inverterade att platser flankerar donator- och mottagarkassetterna i varje ände, vilket driver två oberoende rekombinationshändelser att äga rum samtidigt som det resulterar i kassettutbyte utan integration av plasmid ryggrad. Denna förbättrade konstruktion kringgår integrationen av oönskade sekvenser och utökar tillämpningen av φC31-system till att till exempel omfatta integrering av omärkta DNA-laster genom screening för förlust av en tidigare integrerad fluorescerande markör32.
RMCE uppnåddes först med D. melanogaster32 och tillämpades senare framgångsrikt på icke-modell insekter inklusive An. gambiae9,26,33, Ae. aegypti34, Plutella xylostella34 och B. mori35. Flera dockningslinjer för RMCE har utvecklats och validerats i An. gambiae5,9,26 (tabell 1). Såvitt vi vet har RMCE ännu inte utforskats i andra Anopheles vektorarter.
Hittills har φC31-systemet använts i stor utsträckning i Anopheles myggor för att introducera och studera en mängd olika molekyler, inklusive antimalaria-effektorer19,24,36, komponenter i GAL4/ UAS-systemet för att överuttrycka och slå ner gener för insekticidresistensstudier9,33, regulatoriska element, reportergener5,21,37 och gendrivelement26 ,38.
Detta protokoll beskriver hur man utför 1) platsstyrd integration av en attB-flankerad last och 2) RMCE av en konstruktion flankerad av inverterade attB-platser i genomet av Anopheles dockningslinjer. Detta uppnås med hjälp av två plasmider: en givaren attB-märkt plasmid som bär transgenen av intresse och en hjälpare plasmid som uttrycker φC31 integrase. De stora malariavektorerna An. gambiae och An. stephensi används som specifika exempel, men dessa protokoll är tillämpliga på andra Anopheles arter.
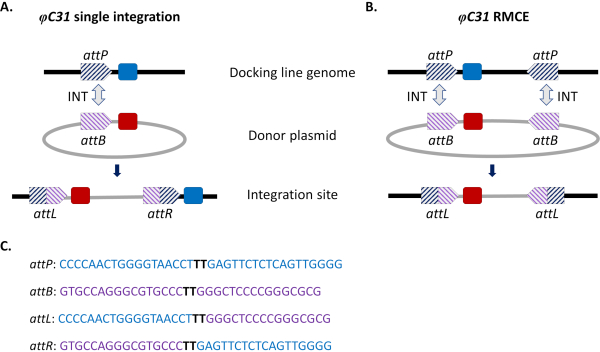
Figur 1. Platsstyrda genomändringar, enkel integrering och rekombinerad kassettutbyte (RMCE) med hjälp av φC31-systemet. φC31 integrase (INT, grå dubbelpil) katalyserar rekombinationen mellan attB-platserna (lila randiga) som finns i en donatorplasmid och attP-platserna (blå randiga) som finns i en mottagningsdockningslinje, vilket resulterar i bildandet av hybridplatser attL och attR. A) Integration uppnås när enstaka attB- och attP-platser kombineras och resulterar i närvaron av två integrerade markörer (blå och röd). B) RMCE inträffar när två attB/P-platser kombineras samtidigt och resulterar i att kassetten byts ut mellan dockningslinjens (blå markör) med den som bärs av donatorplasmiden (röd markör). C) Partiella nukleotidsekvenser av attP (blå) och attB (lila) och hybridplatserna attL/R. Rekombination sker mellan “TT”-kärnsekvenserna markerade i fet svart. Klicka här för att se en större version av den här figuren.
