Hastalıkların sivrisinek vektörlerinin genomlarını güvenilir ve tekrarlanabilir bir şekilde değiştirme yeteneği, genlerin in vivo fonksiyonel doğrulamasını güçlendirmiş ve sıtmayı ileten Anopheles sivrisineklerini hedef alanlar gibi gerçekleştirilebilir genetik vektör kontrol stratejilerine kapı açmıştır1.
Erken sivrisinek genom düzenlemesi sadece transpoze edilebilir eleman (TE) aracılı dönüşüme dayanıyordu ve piggyBac Anopheles2,3,4’te en sık kullanılan transposondu. Bununla birlikte, TE entegrasyonunun rastgele doğası, gen nakavtları (eklemeli mutagenezi) ve transgene ekspresyonu üzerinde önemli konum etkileri gibi istenmeyen değişikliklere yol açabilir5,6,7,8. Birden fazla ekleme, tek eklemelerle transgenik çizgilerin doğrulanmasını ve yalıtılmasını zahmetli hale getiren piggyBac5,9 kullanırken de yaygın bir durumdur. Diğer dezavantajları, bir domuzbac transposase10,11,12 kaynağı sağlarken Anopheles stephensi’nin germline’ında gözlemlendiği gibi potansiyel yeniden hareketlenmelerini ve donör plazmid13,14’ün artan boyutuyla azalan dönüşüm verimliliği ile sınırlı DNA kargo boyutlarını (10-15 kb uzunluğunda) içerir.
Bu sorunları atlatmak için siteye yönelik entegrasyon yaklaşımları getirilmiştir. Sivrisineklerde en yaygın bölgeye yönlendirilen genom modifikasyonu, φC31 sisteminin aracılık ettiğidir (Şekil 1a). Bu, bakteriyofaj φC31 (attP) genomunda ve Streptomices bakteri konağında (attB)15’te doğal olarak meydana gelen iki heterospesifik bağlanma (att) bölgesi arasındaki rekombinasyonu katalizleyen viral bir integrase tarafından yönlendirilir. İki bölgenin yeniden birleşmesi tek yönlüdür ve hibrit bölgelerin (attL ve attR) oluşumuyla sonuçlanır. Bu tür melez bölgelerin yeniden bireşmesi (DNA eksizyona yol açan) sadece aktif bir viral integrazın varlığını değil, aynı zamanda başka bir faj kodlu rekombinasyon faktörünü de gerektirecektir16,17. Böylece, potansiyel istenmeyen yeniden hareketlenme sorununu hafifleten kararlı bir entegrasyon sitesi oluşturulur15. Ayrıca, sistem büyük yüklerin entegrasyonuna izin verir (örneğin, D. melanogaster18’de >100 kb yapı entegrasyonu bildirilmiştir), taşıma kapasitelerini önemli ölçüde artırmaktadır. Entegrasyon, kararlı bir transgenik hat elde etmek için ekleme ve çiftleşme şemasının doğrulanmasını büyük ölçüde basitleştiren önceden tanımlanmış tek bir genomik lokusta gerçekleşir. Son olarak, entegrasyonun meydana gelen doğası, alternatif transgenler aynı çekirgede bulunduğundan ve bu nedenle aynı komşu genomik bağlamda düzenlendiğinden ifadenin normalleşmesine izin verir. Aslında, tekniğin ana uygulamalarından biri, farklı transgenler tarafından aynı lokusa yerleştirildiğinden sonra verilen fenotiplerin doğrudan karşılaştırılmasıdır.
φC31 aracılı entegrasyona ulaşmak iki aşamayı içerir: aşama I, ATTP sahalarını taşıyan transgenik yerleştirme hatlarının oluşturulmasıdır ve aşama II, attB tarafından kuşatılmış bir kargonun yerleştirme hattının genomuna siteye yönelik entegrasyonudur19. Faz I yerleştirme hatlarının oluşturulması, ATTP etiketli yapıların TE aracılı rastgele entegrasyonuna dayanmıştır ve bu nedenle benzersiz, transkripsiyonel olarak aktif ve fitness nötr genomik konumlarda tek bir entegrasyon olayı taşıyan transgenik çizgileri izole etmek ve doğrulamak için ilk zahmetli bir süreci (tek kadın soylar üzerinde güney lekesi ve ters PCR analizleri dahil) içermektedir. Bununla birlikte, An. gambiae19,20,21,22 ve An. stephensi23,24,25’te (Tablo 1) φC31 aracılı tek entegrasyon için birkaç yerleştirme hattı geliştirilmiş ve doğrulanmıştır. Bu çizgilerin her biri, yanaşma bölgesinin genomik konumu ve suşlara özgü genetik arka plan açısından değişir ve bunlardan çok çeşitli yeni transgenik çizgiler oluşturulabilir. Yerleştirme hatları üretmek için TE aracılı entegrasyonların karmaşık doğrulaması artık CRISPR/Cas9 teknolojisi tarafından atlatılabilir26; ancak bu, hedeflenecek nötr loci ve çevresindeki diziler hakkında a priori bilgisine dayanır.
φC31 aracılı entegrasyon, model organizma D. melanogaster27’den Aedes aegypti13,28, Ae. albopictus29, An. gambiae19 ve An. stephensi24’ün yanı sıra Ceratitis capitata30 ve Bombyx mori31 gibi diğer böceklere kadar böcek genom düzenlemesine kapsamlı bir şekilde uygulanmıştır.
φC31 aracılı entegrasyonun bir sınırlaması, özellikle vektör kontrolü için potansiyel alan salınımları göz önüne alınarak, antibiyotik direnci gen belirteçleri ve bakteri kökenli plazmid omurga bileşenleri gibi istenmeyen diziler de dahil olmak üzere tüm ATTB taşıyan donör plazmidinin sivrisinek genomuna entegrasyonudur. Bunu gidermek için, daha önce entegre edilmiş bir transgenik kasetin yeni bir donör DNA’sı ile kesin olarak değiştirilmesini sağlayan standart sistemin, rekombinazi aracılı kaset değişiminin (RMCE) bir modifikasyonu uygulanmıştır (Şekil 1b). Bu, her iki uçta donör ve alıcı kasetleri kuşatan iki ters att bölgesi kullanılarak elde edilir, bu da iki bağımsız rekombinasyon olayının aynı anda gerçekleşmesini sağlar ve plazmid omurgası entegre edilmeden kaset değişimi ile sonuçlanır. Bu geliştirilmiş tasarım, istenmeyen dizilerin entegrasyonunu atlatır ve φC31 sistemlerinin uygulamasını, örneğin daha önce entegre edilmiş bir floresan marker32’nin kaybını tarayarak işaretlenmemiş DNA kargolarının entegrasyonunu içerecek şekilde genişletir.
RMCE ilk olarak D. melanogaster32 ile elde edildi ve daha sonra An. gambiae9,26,33, Ae. aegypti34, Plutella xylostella34 ve B. mori35 dahil olmak üzere model olmayan böceklere başarıyla uygulandı. An. gambiae5,9,26 ‘da RMCE için çeşitli yerleştirme hatları geliştirilmiş ve doğrulanmıştır (Tablo 1). Bilgimize göre, RMCE henüz diğer Anopheles vektör türlerinde araştırılmamıştır.
Bugüne kadar, φC31 sistemi, antimalaria efektörleri19,24,36, GAL4/UAS sisteminin bileşenleri, insektisit direnci çalışmaları için genleri aşırı ifade etmek ve devirmek için çeşitli molekülleri tanıtmak ve incelemek için Anopheles sivrisineklerinde yaygın olarak kullanılmıştır9,33, düzenleyici elementler, muhabir genleri5,21,37 ve gen tahrik elemanları26 ,38.
Bu protokol, 1) ATTB tarafından kuşatılmış bir kargonun saha tarafından yönlendirilen entegrasyonunun ve 2) ters attB siteleri tarafından kuşatılmış bir yapının RMCE’nin Anopheles yerleştirme hatlarının genomuna nasıl gerçekleştirildiğini açıklar. Bu, iki plazmid kullanılarak elde edilir: ilginin transgenesini taşıyan bir donör attB etiketli plazmid ve φC31 integrazını ifade eden bir yardımcı plazmid. Başlıca sıtma vektörleri An. gambiae ve An. stephensi özel örnekler olarak kullanılır, ancak bu protokoller diğer Anopheles türleri için geçerlidir.
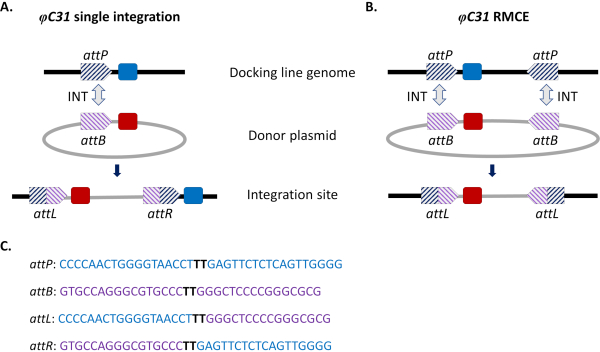
Şekil 1. φC31 sistemi kullanılarak raya yönlendirilmiş genom modifikasyonları, tek entegrasyon ve rekombinoz aracılı kaset değişimi (RMCE). φC31 integraz (INT, gri çift ok), donör plazmidinde bulunan ATTB siteleri (mor çizgili) ile alıcı yerleştirme hattında bulunan attP siteleri (mavi çizgili) arasındaki yeniden birleştirmeyi katalizörler ve bu da attL ve attR’de hibrit sitelerin oluşmasına neden olur. A) Entegrasyon, tek attB ve ATTP siteleri yeniden birleştirildiğinde ve iki entegre işaretleyicinin (mavi ve kırmızı) varlığıyla sonuçlandığında elde edilir. B) RMCE, iki attB/P bölgesi aynı anda yeniden birleşince ortaya çıkar ve kasetin yerleştirme hattının att bölgeleri (mavi işaretleyici) arasında donör plazmid (kırmızı işaretleyici) tarafından taşınanla değiştirilmesiyle sonuçlanır. C) AttP (mavi) ve attB (mor) ve hibrid siteler attL / R kısmi nükleotid dizileri. Yeniden birleştirme, koyu siyah renkle vurgulanan ‘TT’ çekirdek dizileri arasında gerçekleşir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
