एकल-अणु Förster अनुनाद ऊर्जा हस्तांतरण (smFRET) एक ऐसी तकनीक है जो व्यक्तिगत अणुओं के स्तर पर दो रंगों-एक दाता और एक स्वीकर्ता के बीच FRET दक्षता को मापता है। FRET दो रंगों के अतिव्यापी ऊर्जा स्पेक्ट्रा से उत्पन्न होने वाली एक फोटोफिजिकल प्रक्रिया है: दाता एक विशिष्ट तरंग दैर्ध्य के प्रकाश से उत्साहित होता है और स्वीकर्ता को गैर-विकिरणीय रूप से ऊर्जा स्थानांतरित करता है, जिसके परिणामस्वरूप स्वीकर्ता से उत्सर्जन होता है। इस हस्तांतरण की दक्षता दो रंगों के बीच की दूरी की छठी शक्ति के व्युत्क्रमानुपाती है, इसलिए स्थानांतरण दक्षता दूरी 1 के साथ भिन्न होती है। इस प्रकार, इस FRET दक्षता का उपयोग अणु (ओं) 2 के बारे में स्थानिक जानकारी निर्धारित करने के लिए किया जा सकता है, जिसमें रंजक जुड़े हुए हैं, 3-10 एनएम की सीमा के भीतर। यह पैमाने पर, और तथ्य यह है कि FRET दक्षता में परिवर्तन Angstrom आणविक आंदोलनों 3 के प्रति संवेदनशील हैं, तकनीक को बायोमोलेक्यूल्स के बारे में संरचनात्मक जानकारी की जांच करने के लिए अच्छी तरह से अनुकूल बनाता है- जैसे न्यूक्लिक एसिड और प्रोटीन-पहनावा औसत की जटिलताओं के बिना 4,5,6। जबकि सापेक्ष FRET दक्षताओं में परिवर्तन का उपयोग बायोमोलेक्यूलर इंटरैक्शन और संरचनात्मक गतिशीलता की निगरानी के लिए किया जा सकता है, प्रोटीन (un) तह, प्रतिलेखन, और डीएनए प्रतिकृति और मरम्मत जैसी प्रमुख सेलुलर प्रक्रियाओं पर प्रकाश डाला जा सकता है, पूर्ण FRET दक्षता का उपयोग बायोमोलेक्यूलर संरचना निर्धारण के लिए सटीक दूरी निर्धारित करने के लिए किया गया है7,8,9,10,11 , क्रिस्टलीकरण या ठंड की आवश्यकता पर काबू पाने के रूप में कुछ अन्य संरचनात्मक तरीकों के लिए आवश्यक है4,12.
smFRET प्रयोगों सबसे अधिक दो रूपों, confocal या कुल आंतरिक प्रतिबिंब प्रतिदीप्ति (TIRF) माइक्रोस्कोपी ले लो. दोनों दृष्टिकोणों के बीच बायोमोलेक्यूल्स की आणविक गतिशीलता की जांच आमतौर पर पिको से मिलीसेकंड (कॉन्फोकल, स्वतंत्र रूप से अलग अणुओं) से मिलीसेकंड से लेकर घंटों तक (टीआईआरएफ, सतह स्थिर अणुओं) तक टाइमस्केल पर की जा सकती है। यह प्रत्येक तकनीक में शामिल विभिन्न सेटअपों के कारण है। TIRF माइक्रोस्कोपी में, अणुओं को स्लाइड की सतह पर स्थिर किया जाता है और एक तरंग (चित्रा 1 ए) द्वारा उत्तेजित किया जाता है। यहां, हालांकि, फोकस कॉन्फोकल माइक्रोस्कोपी पर है क्योंकि यह smfBox का प्रारूप है। Confocal माइक्रोस्कोपी में, अणुओं को स्थिर नहीं किया जाता है और इसके बजाय स्वतंत्र रूप से confocal मात्रा (~ 1 fL) के माध्यम से ब्राउनियन गति के माध्यम से फैलाया जाता है, जो समाधान के भीतर कुछ निर्दिष्ट गहराई पर एक स्थान पर एक उच्च संख्यात्मक एपर्चर लेंस के माध्यम से एक लेजर बीम पर ध्यान केंद्रित करके बनता है (चित्रा 1 बी)। परिणामी उत्सर्जन को एक ही एपर्चर के माध्यम से वापस केंद्रित किया जाता है और एक डाइक्रोइक दर्पण के माध्यम से फ़िल्टर किया जाता है (पूर्ण योजनाबद्ध के लिए चित्रा 1 सी )। यह तब किसी भी आउट-ऑफ-फोकस प्रकाश को हटाने के लिए और एक हिमस्खलन फोटोडायोड (एपीडी) पर पिनहोल के माध्यम से केंद्रित होता है। जब एपीडी एक फोटॉन का पता लगाता है, तो यह एक टीटीएल पल्स आउटपुट करता है, जिसका समय पिकोसेकंड रिज़ॉल्यूशन तक रिकॉर्ड किया जा सकता है। कॉन्फोकल वॉल्यूम के आसपास के क्षेत्र के भीतर इन स्वतंत्र रूप से अलग अणुओं का अवलोकन समय आमतौर पर मिलीसेकंड के क्रम के भीतर होता है।
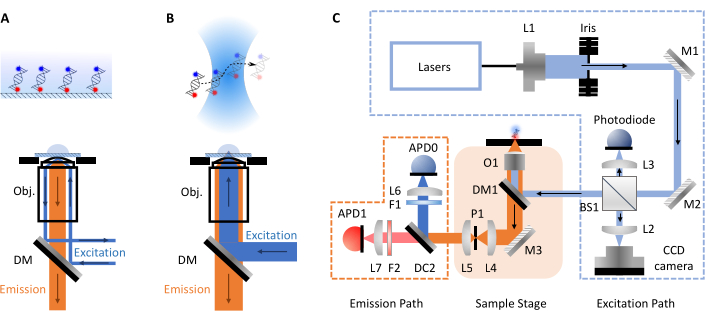
चित्र1: माइक्रोस्कोपी और smfBox सेटअप के सिद्धांतों को दिखाने वाली योजनाबद्ध योजनाएं। (A) कुल आंतरिक प्रतिबिंब प्रतिदीप्ति (TIRF) माइक्रोस्कोपी सिद्धांत: उत्तेजना प्रकाश को उद्देश्य (Obj.) के किनारे में निर्देशित किया जाता है और कवरस्लिप-बफर इंटरफ़ेस पर कुल आंतरिक प्रतिबिंब से गुजरता है जो सतह से जुड़े अणुओं को उत्तेजित करने के लिए एक तेजी से क्षयकारी evanescence क्षेत्र उत्पन्न करता है। (बी) कॉन्फोकल माइक्रोस्कोपी: स्वतंत्र रूप से विवर्तन करने वाले अणु नमूने में केंद्रित एक निकट विवर्तन-सीमित स्थान से उत्साहित होते हैं। (सी) इस प्रोटोकॉल में उपयोग किया जाने वाला smfBox सेटअप, सभी प्रमुख घटकों को दर्शाता है: हिमस्खलन फोटोडायोड (एपीडी), बीम स्प्लिटर (बीएस), डाइक्रोइक दर्पण (डीएम), फिल्टर (एफ), दर्पण (एम), उद्देश्य (ओ) और पिनहोल (पी)। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहाँ क्लिक करें.
हाल ही में, smFRET तकनीकों में दो रंग उत्तेजना शामिल हैं, जहां दाता और स्वीकर्ता उत्तेजना तरंग दैर्ध्य से मेल खाने वाले लेजर को वैकल्पिक किया जाता है5। यह दो तरीकों में से एक में किया जा सकता है, पहला KHz टाइमस्केल पर निरंतर तरंग लेजर को मॉड्युलेट करके, जिसे वैकल्पिक लेजर उत्तेजना (एलेक्स) 13,14 के रूप में जाना जाता है। दूसरी विधि MHz टाइमस्केल पर तेजी से दालों interleaves; यह nanosecond-ALEX15 या स्पंदित interleaved उत्तेजना (PIE)16 है. इन सभी दृष्टिकोणों में, स्वीकर्ता लेजर से जानकारी तथाकथित स्टोइकोमेट्री की गणना की ओर ले जाती है, जो कम FRET दक्षता वाले अणुओं और स्वीकर्ता की कमी वाले अणुओं के बीच भेदभाव कर सकती है (या तो अपूर्ण लेबलिंग या फोटोब्लीचिंग के माध्यम से)। पीआईई / एनएस-एलेक्स का उपयोग करना अतिरिक्त रूप से एकल-अणु स्तर पर फ्लोरोसेंट जीवनकाल तक पहुंच प्रदान करता है, और ध्रुवीकरण प्रकाशिकी के साथ युग्मित होने पर अनिसोट्रॉपी को मापा जा सकता है। माप के इस संयोजन को मल्टीपैरामीटर प्रतिदीप्ति का पता लगाने (MFD)9 के रूप में जाना जाता है।
SMFRET के कई फायदों के बावजूद, वाणिज्यिक उपकरणों की उच्च लागत और सरल, स्व-निर्माण विकल्पों की कमी के कारण विशेषज्ञ प्रयोगशालाओं के बाहर इसका व्यापक रूप से उपयोग नहीं किया जाता है। कम लागत वाले ओपनसोर्स माइक्रोस्कोपी के विकास की दिशा में एक बढ़ती प्रवृत्ति हो रही है और अन्य प्लेटफार्म हाल ही में उभरे हैं, जिनमें प्लैंकटनस्कोप 17, ओपनफ्लेक्स्योर माइक्रोस्कोप 18, फ्लेक्सिस्कोप 19, miCube20, liteTIRF21 और Squid22 शामिल हैं। इसमें अध्ययन smfBox का उपयोग करने के लिए प्रोटोकॉल का वर्णन करता है, एक हाल ही में विकसित लागत प्रभावी confocal सेट-अप स्वतंत्र रूप से एकल अणुओं diffusing पर दो रंगों के बीच FRET दक्षता को मापने में सक्षम है। विस्तृत निर्माण निर्देश और सभी आवश्यक परिचालन सॉफ़्टवेयर स्वतंत्र रूप से उपलब्ध हैं: https://craggslab.github.io/smfBox/ 23। SmfBox की ऑप्टिकल व्यवस्था को सस्ती और व्यापक रूप से सुलभ निर्माताओं से खरीदे गए आसानी से उपलब्ध घटकों से इकट्ठा किया जाता है, जबकि माइक्रोस्कोप बॉडी (एक मानक कॉन्फोकल सेट-अप में अधिकांश खर्च के लिए जिम्मेदार) को एक कस्टम लाइट-टाइट एनोडाइज्ड-एल्यूमीनियम बॉक्स द्वारा प्रतिस्थापित किया गया है (परिवेश प्रकाश परिस्थितियों के तहत माप करने की अनुमति देता है)। इस बॉक्स में प्रमुख ऑप्टिकल घटक हैं, जिनमें उत्तेजना डाइक्रोइक, उद्देश्य और पिनहोल, और एक यांत्रिक लेजर इंटरलॉक शामिल हैं, जो कक्षा I लेजर उत्पाद के रूप में अपने सुरक्षित संचालन को सक्षम करते हैं (एक पूर्ण योजनाबद्ध के लिए चित्रा 1 सी देखें)। SmfBox एलेक्स का उपयोग करता है डाई stoichiometry मान्य करने के लिए और सटीक FRET सुधार कारकों को निर्धारित करने के लिए. यह कस्टम-लिखित, ओपन-सोर्स सॉफ़्टवेयर (smOTTER) का उपयोग करके संचालित किया जाता है, जो डेटा अधिग्रहण के सभी पहलुओं को नियंत्रित करता है और कई तृतीय-पक्ष विश्लेषण उपकरणों के साथ संगत ओपन-सोर्स फोटॉन-HDF5 format24 में डेटा आउटपुट करता है। SmfBox और अधिग्रहण और डेटा विश्लेषण प्रोटोकॉल हाल ही में एक बहु-प्रयोगशाला अंधा अध्ययन 25 में >20 अन्य उपकरणों (confocal और TIRF दोनों) के खिलाफ परीक्षण किया गया था। प्राप्त FRET दक्षता अन्य सभी उपकरणों के साथ उत्कृष्ट समझौते में थे, smfBox व्यावसायिक रूप से उपलब्ध सेटअप की कीमत का केवल एक अंश लागत के बावजूद।
यहां, एक चरण-दर-चरण प्रोटोकॉल को प्राप्त करने और विश्लेषण करने के लिए रेखांकित किया गया है, जो कि smfBox का उपयोग करके डीएनए डुप्लेक्स को स्वतंत्र रूप से अलग करने पर सटीक, पूर्ण FRET दक्षताओं को प्राप्त करने और विश्लेषण करने के लिए, सभी तरह से स्विच ऑन से, संरेखण और ध्यान केंद्रित करने के माध्यम से, डेटा संग्रह और विश्लेषण के लिए। यहां उपयोग किए जाने वाले नमूने तीन डुप्लेक्स डीएनए हैं (उच्च, मध्य और निम्न-FRET दक्षताओं को प्रदर्शित करते हुए, तालिका 1 देखें) जो दुनिया भर में अंधे अध्ययन 25 में मूल्यांकन किए गए थे; हालांकि, विधि प्रोटीन और अन्य न्यूक्लिक एसिड सहित कई आणविक प्रणालियों के लिए अनुकूलहै। आशा है कि इस तरह के एक विस्तृत प्रोटोकॉल, smfBox23 के लिए पहले से ही मौजूदा निर्माण निर्देशों के साथ, इस शक्तिशाली तकनीक को प्रयोगशालाओं की एक विस्तृत श्रृंखला के लिए और भी अधिक सुलभ बनाने में मदद करेगा।
