In this study, the purified soluble APN protein (2.12 mg/mL) was used for mouse immunization. Mice immunized with the APN protein four times at 14-day intervals exhibited a higher antibody titer against APN in their sera. Although 14 hybridomas were obtained using the fusion experiments, only 9 hybridomas survived the three continuous freeze-thaw cycles, resulting in 9 stable clones that secreted antibodies against APN. All these cells are round, bright, and clear (Figure 1). The purified mAbs possessing heavy and light chains (50 kDa and 25 kDa, respectively) were confirmed by SDS-PAGE and found in the purified ascites (Figure 2). The titers of these anti-APN mAbs in culture supernatants and ascites are shown in Table 2.
The result of mouse mAbs isotyping revealed that mAbs derived from clones 5B31, 5B36, 3C48, 5C51, and 6C56 possessed IgG2b subclasses, while APN-2A20 was an IgG2a κappa- (κ) type antibody, and mAbs APN-3FD9, -3F10, and -10F3 belonged to IgM type and processed κ light chains (Table 3). As shown in Table 4, most of these mAbs showed AV values of over 50%, indicating that they targeted different epitopes in the APN, while the APN-5C51 antibody recognized antigenic epitopes similar to those recognized by APN-3C48, -5B31, and -6C56 mAbs.
APN-5B36 showed considerably higher antibody titer compared with those of other mAbs. Therefore, the APN-5B36 VH-VL gene was amplified and ligated into a pET28a (+) or pIRES2-ZsGreen1 vector to construct the recombinant expression plasmids pET28a (+)-rAbs-APN and pIRES2-ZsGreen1-rAbs-APN, respectively (Figure 3). The antibodies expressed by both pET28a (+)-rAbs-APN-BL21 (DE3) and pIRES2-ZsGreen1-rAbs-APN-CHO cells were purified and analyzed using ELISA and IFA assays. However, as shown in Figure 4, only the antibody expressed in the supernatant of pIRES2-ZsGreen1-rAbs-APN-CHO cells recognized the APN protein, as did hybridoma-derived mAbs. This recombinant antibody consisted of IgG2b heavy chains and lambda light chains, and showed a titer of 2.56 × 105 as determined using ELISA. The binding of APN-5B36 mAbs to APN proteins reached an equilibrium earlier than rAbs did (Figure 5), showing KD value of (4.232±0.475) × 10-9 and (2.201±0.367) × 10-8 mol/L, respectively.
| Primer | Sequence (5’-3’) | ||
| VH-VL-F | CCGGGTGGGCCGGATAGACMGATGGGGCTG | ||
| VH-VL-R | CCGGCCACATAGGCCCCACTTGACATTGATGT | ||
| pET28a (+)-F | TCCACCAGTCATGCTAGCCATAACAACGGTCGTGATTCGA | ||
| pET28a (+)-R | CTGGTGCCGCGCGGCAGCCAGTGGGATACCCGTATTACCC | ||
| pIRES2-ZsGreen1-F | CGACGGTACCGCGGGCCCGGTAACAACGGTCGTGATTCGA | ||
| pIRES2-ZsGreen1-R | GGGGGGGAGGGAGAGGGGCGGTGGGATACCCGTATTACCC | ||
Table 1. The specific primers used in this study.
| Cells | Titers of supernatants (U/mL) | Titers of ascites (U/mL) |
| 2A20 | 0.64×104 | 3.20×105 |
| 5B31 | 1.28×104 | 1.60×105 |
| 5B36 | 0.64×104 | 1.28×106 |
| 3C48 | 0.16×104 | 0.80×105 |
| 5C51 | 0.16×104 | 0.80×104 |
| 6C56 | 0.80×103 | 0.80×104 |
| 3FD9 | 0.80×103 | 0.80×104 |
| 3F10 | 0.16×104 | 0.16×105 |
| 10F3 | 0.80×103 | 0.32×105 |
Table 2. The ELISA titers of APN mAbs.
| Ig | IgA | IgM | IgG1 | IgG2a | IgG2b | IgG3 | Kappa | Lambda | Summary | |
| 2A20 | 1.735 | 0.023 | 0.011 | 0.006 | 0.903 | 0.044 | 0.015 | 0.137 | 0.073 | IgG2a, Kappa |
| 5B31 | 1.199 | 0.006 | 0.003 | 0.005 | 0.005 | 1.731 | 0.004 | 0.004 | 0.413 | IgG2b, Lambda |
| 5B36 | 1.652 | 0.012 | 0.013 | 0.01 | 0.008 | 2.41 | 0.002 | 0.003 | 0.707 | IgG2b, Lambda |
| 3C48 | 0.951 | 0.063 | 0.068 | 0.104 | 0.062 | 1.785 | 0.059 | 0.065 | 0.51 | IgG2b, Lambda |
| 5C51 | 1.064 | 0.008 | 0.007 | 0.008 | 0.008 | 1.87 | 0.004 | 0.004 | 0.415 | IgG2b, Lambda |
| 6C56 | 0.78 | 0.062 | 0.06 | 0.063 | 0.063 | 1.516 | 0.062 | 0.061 | 0.387 | IgG2b, Lambda |
| 3FD9 | 1.474 | 0.007 | 1.678 | 0.003 | 0.016 | 0.081 | 0.002 | 0.519 | 0.059 | IgM, Kappa |
| 3F10 | 1.21 | 0.002 | 1.454 | 0.009 | 0.008 | 0.054 | 0.003 | 0.414 | 0.096 | IgM, Kappa |
| 10F3 | 1.179 | 0.058 | 1.562 | 0.152 | 0.131 | 0.179 | 0.044 | 0.359 | 0.049 | IgM, Kappa |
Table 3. Isotypes of hybridoma-derived APN mAbs.
| mAbs | AV (100 %) | ||||||||
| 2A20 | 5B31 | 5B36 | 3C48 | 5C51 | 6C56 | 3FD9 | 3F10 | 10F3 | |
| 2A20 | – | 0.601 | 0.905 | 0.889 | 0.804 | 0.884 | 1.009 | 1.047 | 0.914 |
| 5B31 | 0.601 | – | 0.871 | 0.754 | 0.464 | 0.694 | 0.613 | 0.88 | 0.989 |
| 5B36 | 0.905 | 0.871 | – | 0.794 | 0.684 | 0.934 | 0.91 | 1.07 | 0.959 |
| 3C48 | 0.889 | 0.754 | 0.794 | – | 0.461 | 0.709 | 0.428 | 1 | 0.787 |
| 5C51 | 0.804 | 0.464 | 0.684 | 0.461 | – | 0.301 | 0.601 | 0.594 | 0.852 |
| 6C56 | 0.884 | 0.694 | 0.934 | 0.709 | 0.301 | – | 1.216 | 0.583 | 0.389 |
| 3FD9 | 1.009 | 0.613 | 0.91 | 0.428 | 0.601 | 1.216 | – | 1.737 | 0.744 |
| 3F10 | 1.047 | 0.88 | 1.07 | 1 | 0.594 | 0.583 | 1.737 | – | 0.682 |
| 10F3 | 0.914 | 0.989 | 0.959 | 0.787 | 0.852 | 0.389 | 0.744 | 0.682 | – |
Table 4. Discrimination of antigen-epitope specificity of APN-specific mAbs. AV values greater than 0.5 indicate that these two mAbs recognize different antigenic sites; AV values less than 0.5 indicate that these two mAbs recognize a similar antigenic site.
Figure 1. Image of hybridomas. Under microscopic analysis, hybridomas are round, bright, and clear. Please click here to view a larger version of this figure.
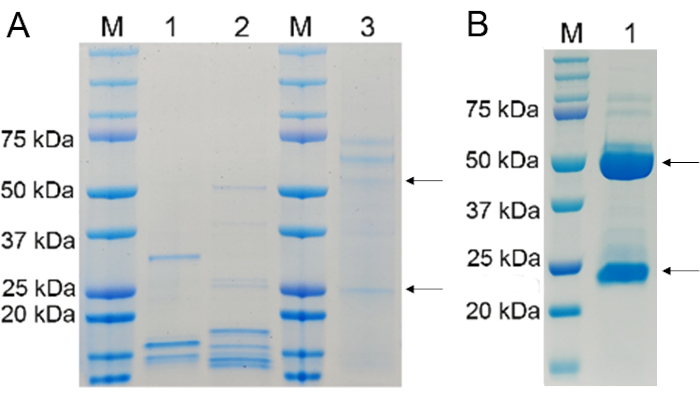
Figure 2. Recombinant antibody expression levels and ascites analyzed using SDS-PAGE. (A) Lane M, protein marker; lane 1, purified pET28a (+)-rAbs-APN-BL21 (DE3) lysate; lane 2, pET28a (+)-rAbs-APN-BL21 (DE3) supernatant; lane 3, ascites fluid purified by 33% (NH4)2SO4 precipitation. (B) Lane M, protein marker; lane 1, ascites fluid purified using protein A agarose. In this assay, 3-5 µg of total protein was loaded into each lane of the gel. Please click here to view a larger version of this figure.
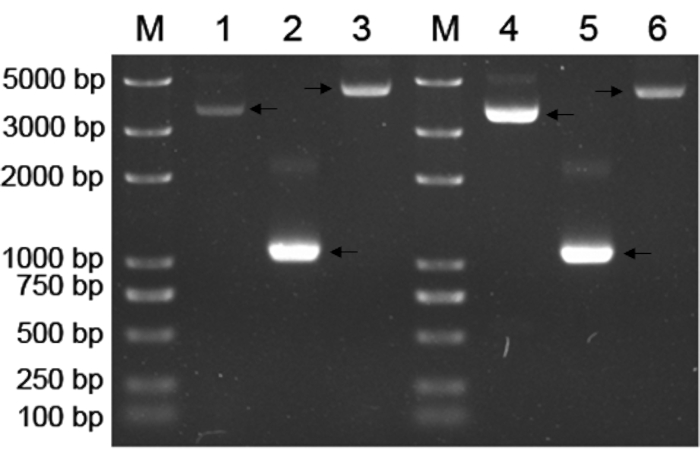
Figure 3. Recombinant expression plasmids pET28a (+)-rAbs-APN and pIRES2-ZsGreen1-rAbs-APN analyzed using agarose gel electrophoresis. Lane M, Trans 2K plus DNA marker; lane 1, pET28a (+) vector (5369 bp); lane 2 and 5, VH-VL gene combined with APN-5B36 leader sequence; lane 3, pET28a (+)-rAbs-APN plasmid expressed in BL21 (DE3) E. coli; lane 4, pIRES2-ZsGreen1 vector (5283 bp); lane 6, pIRES2-ZsGreen1-rAbs-APN plasmid expressed in DH5α E. coli. Please click here to view a larger version of this figure.
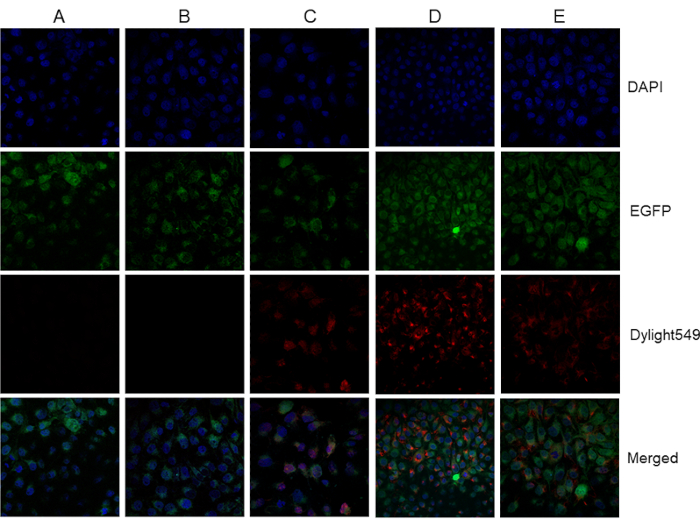
Figure 4. Expression of recombinant antibody protein and ascites analyzed using indirect immunofluorescence. pEGFP-C1-APN-IPEC-J2 cells (green fluorescence) stably expressing APN were treated with (A) PBS, used as control treatment; (B) purified protein expressed by pET28a (+)-rAbs-APN-BL21 (DE3); (C) APN polyclonal antibody (1:500); (D) purified ascites fluid (1:500); E) purified supernatant obtained from pIRES2-ZsGreen1-rAbs-APN-CHO cells (1:500). DAPI was used as nuclear counterstain in confocal microscopy. The cells incubated with the DyLight 549-conjugated goat anti-mouse IgG secondary antibody (1:200) and treated with purified ascites and purified supernatant from pIRES2-ZsGreen1-rAbs-APN-CHO cells showed a robust red-fluorescence signal indicative as the APN polyclonal antibody did. Please click here to view a larger version of this figure.
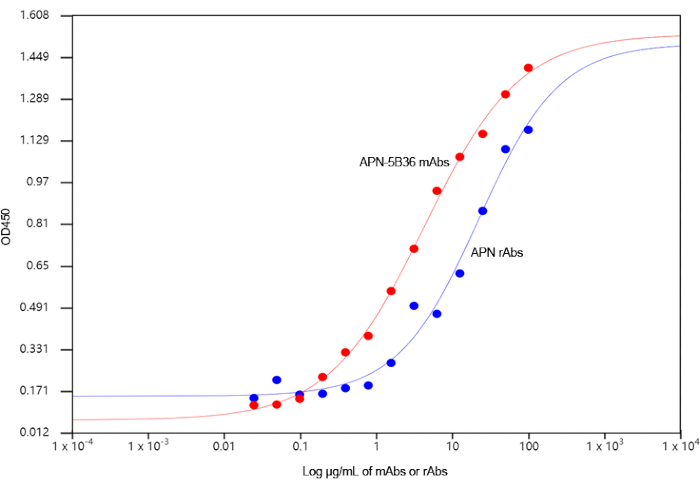
Figure 5. Determination of antibody relative binding affinities using the ELISA22. Absorption of samples containing APN-5B36 mAbs or rAbs, in the absence and presence of APN protein, was measured at the wavelength of 450 nm. Binding curve was plotted using a four-parameter logistic curve fit; x-axis shows the logarithmic concentration of antibodies, and y-axis shows the absorbance. Please click here to view a larger version of this figure.
