Data obtained using this protocol were originally published in Blood in 20169.
According to the protocol, the cells were seeded in either liquid or methylcellulose hydrogel medium. Cells in liquid medium have all sedimented at the bottom of the well, in contact with the stiff plastic surface and sometime with other cells. In contrast, cells embedded in methylcellulose hydrogel are distributed homogeneously in the gel and are isolated from neighboring cells (Figure 3A). Methylcellulose gel at a final concentration of 2% very slightly increases the mean megakaryocyte diameter compared to the liquid culture (Figure 3B), in accordance with the higher reported ploidy9. By contrast, increasing methylcellulose concentration by 0.5% impairs megakaryocyte differentiation as shown by a smaller mean diameter (Figure 3B).
A noticeable difference in megakaryocyte ultrastructure is observed between megakaryocytes differentiated in liquid culture and those differentiated in vivo within the bone marrow. A characteristic feature of mature megakaryocytes is a complex intracytoplasmic membrane network, the DMS (Demarcation Membrane System) which serves as a reservoir for the membrane of the future platelets. In mature megakaryocytes the DMS organizes to form intertwined membrane sheets which occupy most of the cytoplasm. By transmission electron microscopy (TEM), they appear to be closely apposed and delineate cytoplasmic territories (Figure 3C upper panel) (for the TEM procedure, see the paper method "In situ exploration of the major steps of megakaryopoiesis using transmission electron microscopy" in this same JOVE Issue). In liquid culture, DMS membranes have mostly the appearance of small round, oval, or elongated vesicles without delimitation of cytoplasmic territories (Figure 3C middle panel). By contrast, 2% methylcellulose culture promotes the organization of the DMS in a majority of megakaryocytes, with membranes closely apposed and delimiting cytoplasmic territories, resembling the one in situ (Figure 3C lower panel). This result indicates that the 2% methylcellulose hydrogel culture allows for better megakaryocyte differentiation due to the mechanical constraints of the environmental medium.
After cell transfer into liquid medium at day 3, megakaryocytes begin to extend proplatelets after 4 h 9. Figure 4 shows the quantification of the proportion of megakaryocytes having extended proplatelets 24 h after resuspension in liquid milieu. Ten images were randomly acquired per well, using bright field microscopy and the 20× objective (Figure 4A). The quantification was performed blindly and manually using the cell counter plugin on Fiji (ImageJ) (Figure 4B). Because these are primary cell cultures, there is an inter-experiment variability but the protocol remains robust and offers a good reproducibility. In the liquid pre-culture condition, the proplatelet proportion should be between 10% and 20% whereas this proportion is doubled for the hydrogel pre-culture.
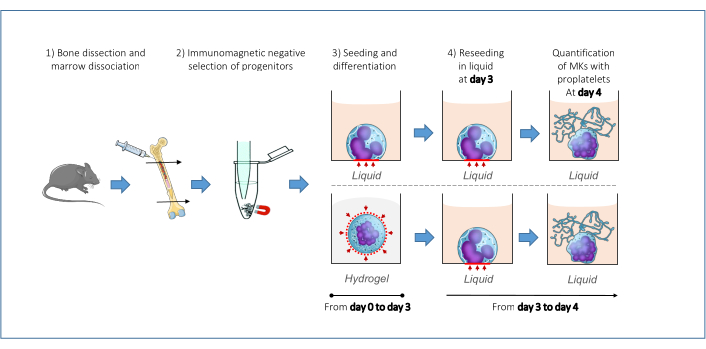
Figure 1. Schematic representation of the whole process. Bones are dissected out, marrow is flushed out and cells mechanically dissociated. Stem and progenitor cells of interest (Lin- cells) are isolated by an immunomagnetic negative sorting procedure and seeded in either liquid or hydrogel medium (day 0). At day 3 of culture (which in total represents a duration of 4 days), both conditions are resuspended in separate fresh liquid culture milieu. This second culturing step is carried out from day 3 until day 4 of culture. The proportion of MKs extending proplatelets is measured at day 4 of culture. For visual clarity, one cell is schematized per well. The blue circle is depicting a single cell with its nucleus in purple. In the final step, both MKs are represented with proplatelet. The proportion of MKs forming proplatelets varies depending on liquid or methylcellulose pre-culture. Please click here to view a larger version of this figure.

Figure 2. Cell embedding in methylcellulose hydrogel. After pre coating the syringes wall, (A) draw the appropriate volume of methylcellulose; (B, C) disconnect the needle and screw a connector onto the syringe; (D) push the methylcellulose halfway through the connector and attach a second syringe; (E) distribute equally the methylcellulose between the two syringes and disconnect them; (F) add the cell suspension to the syringe bearing the connector; (G) reconnect the two syringes; (H) homogenize by pushing the whole volume from one syringe to the other a few times; (I) seed the cells by expelling the whole volume into a culture dish. Please click here to view a larger version of this figure.
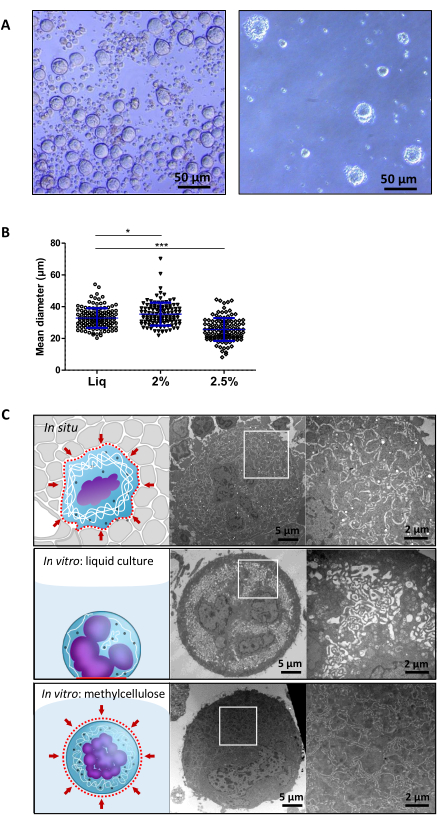
Figure 3. Megakaryocyte characteristics according to culture condition. (A) Representative images of megakaryocytes at day 3 of culture in liquid (left panel) or 2% methylcellulose hydrogel medium (right panel). Scale bar = 50 µm (B) Mean diameter of megakaryocytes grown in liquid medium, or in 2% or 2.5% methylcellulose hydrogel. Results are expressed as the mean ± SD in 3 independent cultures, with a total of at least 100 megakaryocytes examined. *, P<0.05, ***P < 0.0001, using 1-way analysis of variance (ANOVA) with Bonferroni's multiple comparison test. (graph adapted from Aguilar et al. 2016) (C) Schematic view (left) and representative electronic microscopy images (middle) of murine megakaryocytes; right panels, close up views from the white squares (scale bar = 5 µm for the middle electronic microscopy images and 2 µm for close up views). Upper panels are in situ megakaryocytes, middle represents megakaryocytes grown in vitro in liquid culture and lower panels are megakaryocytes grown in 3D methylcellulose hydrogel. These data were originally published in Blood Journal, DOI10.1007/978-1-4939-8585-2_95. Please click here to view a larger version of this figure.
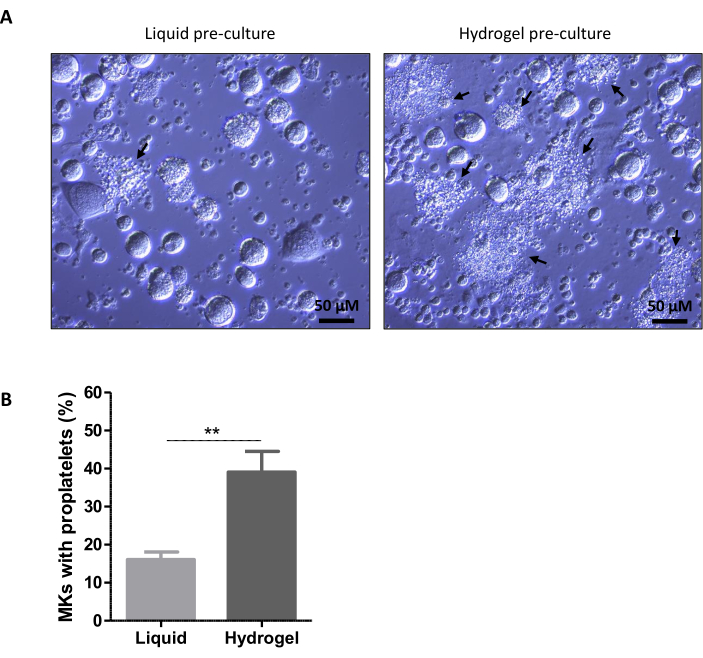
Figure 4. Representative results of proplatelet quantification. (A) representative images of megakaryocytes at day 4 of culture. Cells were incubated three days in liquid (left) or 2% methylcellulose hydrogel medium (right) followed by one day of resuspension in liquid medium. Black arrows indicate megakaryocytes extending proplatelets. (Scale bar = 50 µm). (B) Representative quantification data of proplatelet formation. Proplatelet formation quantified at day 4 for megakaryocytes previously pre-cultured from day 0 to day 3 in liquid or 2% methylcellulose hydrogel medium. Results are expressed as the% of megakaryocytes extending proplatelets (mean ± SD) and are from 3 independent experiments, with a total number of megakaryocytes examined per condition >750 (t-test, p = 0.0023). The mean proportion of megakaryocytes extending proplatelets is 16% in liquid condition and 39% for the methylcellulose hydrogel pre-culture. This result corresponds to the previously demonstrated and published effect of hydrogel pre-culture that increases proplatelet formation compared to liquid condition. Please click here to view a larger version of this figure.
