В течение нескольких лет ооцит дрозофилы появился в качестве модельной системы для изучения ядерной миграции. Ооцит дрозофилы развивается в многоклеточной структуре, называемой камерой яйцеклетки. Яйцеклетки охватывают 16 половых клеток (15 клеток-кормилиц и яйцеклетку), окруженных эпителиальным слоем фолликулярных соматических клеток. Развитие яичной камеры было подразделено на 14 стадий(рисунок 1А),в ходе которых яйцеклетка будет расти и накапливать запасы, необходимые для раннего развития эмбриона. При развитии, при реорганизации микротрубочек и асимметричном переносе материнских детерминант, ооцит поляризуется по антеродорсальной и дорсо-вентральной осям. Эти оси определяют последующие оси полярности эмбриона и взрослого человека, возникающие в результате оплодотворения этой яйцеклетки1. Во время оогенеза ядро принимает асимметричное положение в ооците. На стадии 6 ядро центрируется в клетке. После еще не идентифицированного сигнала, излучаемого задними фолликулярными клетками, который принимается яйцеклеткой, ядро мигрирует к пересечению передней и боковой плазматических мембран на стадии 7(Рисунок 1B)2,3. Это асимметричное положение требуется для того, чтобы вызвать определение дорсо-вентральной оси.
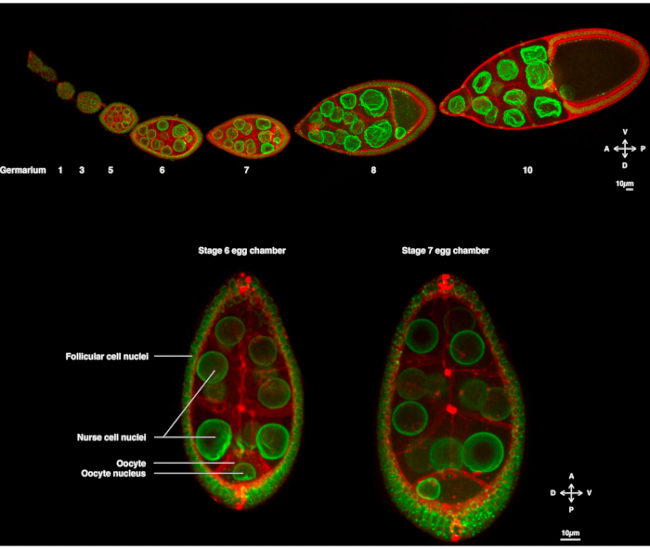
Рисунок 1:Камеры яиц Drosophila melanogaster. ( A )Фиксированныйовариол от трансгенных мух, экспрессивляющих Fs(2)Ket-GFP, который маркирует ядерные оболочки и ubi-PH-RFP, который маркирует плазматические мембраны. Овариол состоит из развивающихся яйцеклеток на разных стадиях. Созревание увеличивается вдоль передне-задней оси с гермарием на переднем кончике (слева), где находится зародышевая стволовая клетка, и более старой стадией на заднем кончике (справа). (B)Z-проекция живой яйцеклетки методом вращающейся дисковой конфокальной микроскопии на 6 стадии оогенеза (слева), при которой ядро центрируется в ооците. Ядро будет мигрировать, чтобы принять асимметричное положение на стадии 7 (справа) в контакте с передней плазматической мембраной (между яйцеклеткой и клеткой медсестры) и боковой плазматической мембраной (между яйцеклеткой и фолликулярными клетками). Это положение будет стимулировать определение дорсальной стороны и, таким образом, дорсо-вентральной оси яйцеклеточной камеры. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
В течение многих десятилетий эта ядерная миграция изучалась на фиксированных тканях путем иммуноокрашивания. Такой подход позволил, в частности, продемонстрировать, что этот процесс зависит от плотной сети микротрубочек4,5. Совсем недавно мы разработали протокол, предлагающий условия, совместимые с живой визуализацией ооцита в течение нескольких часов, что позволяет динамически изучать этот процесс6.
Следовательно, впервые мы смогли описать, что ядро имеет предпочтительные и характерные траектории во время своей миграции, одну вдоль передней плазматической мембраны (APM), а другую вдоль боковой плазматической мембраны (LPM) ооцита(Рисунок 2). Эти последние результаты подчеркивают важность протоколов визуализации в реальном времени при изучении динамических процессов, таких как ядерная миграция.
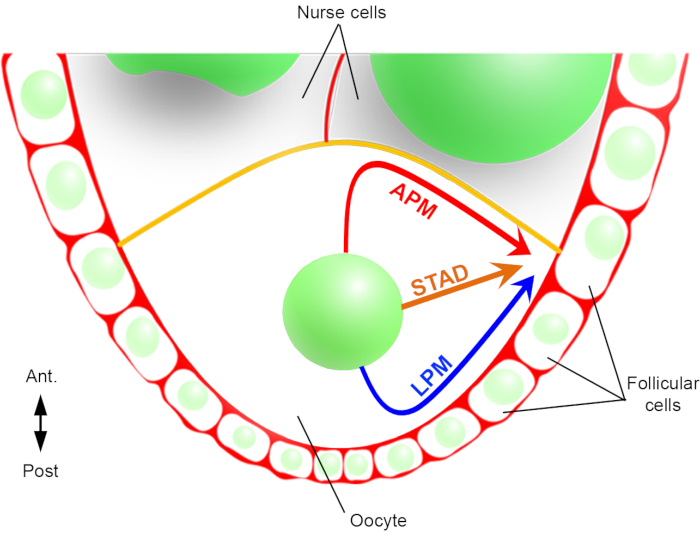
Рисунок 2:Схематическое представление различных путей миграции ядра. На 6 стадии оогенеза яйцеклетка представляет собой крупную клетку с центральным ядром. На этом этапе передне-задняя ось полярности устанавливается с задней/латеральной плазматической мембраной ооцита, контактиим с фолликулярными клетками, а передняя плазматическая мембрана (желтого цвета) контактирует с клетками медсестры2. Ранее мы сообщали, что ядро может мигрировать либо вдоль передней плазматической мембраны (APM), вдоль боковой плазматической мембраны (LPM), либо через цитоплазму (STAD, прямо в антеродоральную кору)6. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Миграция ядра ооцитов представляет собой явление около 3 ч6,и до сих пор событие, вызывающее начало фактической миграции, неизвестно. Начало миграции также может быть отложено белковыми мутантами, используемыми для изучения этого механизма. Эти неизвестные переменные мотивировали нас получать изображения в течение длительных периодов времени (10-12 ч). Поэтому важно обеспечить, чтобы ооциты оставались живыми. По мере развития яичной камеры она удлиняется вдоль антеро-задней оси от сферической до эллиптической формы. Это удлинение обусловлено вращением фолликулярных клеток, которое происходит от стадии 1 до стадии 8, перпендикулярной антеро-задней оси7. Кроме того, трубчатая оболочка мышц с пульсирующим свойством окружает яйцеклетки камеры. Его физиологическая функция заключается в том, чтобы подталкивать развивающиеся фолликулы к яйцеводу непрерывно8. Чтобы ограничить движения, которые вызывают колебания яичных камер после их рассечения, мы разработали наблюдательную микрокамеру высотой 150 мкм(рисунок 3А). Эта высота незначительно превышает размер фолликула на стадиях 10 и 11. Это значительно ограничивает вертикальные движения образца, сохраняя при этом вращение яйцеклеточной камеры, что приводит к ограниченным дефектам в развитии фолликулов. Затем мы выполняем живую визуализацию в течение 12 часов на расчлененных яйцеклетках с помощью многопозиционных покадровых захватов с использованием конфокального микроскопа со вращающимся диском. Здесь мы описываем наш протокол изучения миграции ядер ооцитов между стадиями 6 и 7.
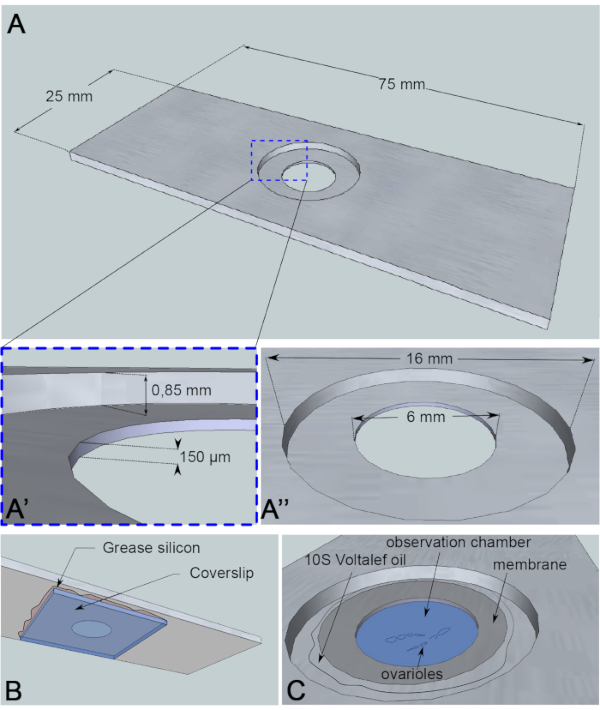
Рисунок 3:Схематическое изображение камеры наблюдения. (A) (Вид сверху) Точные размеры алюминиевой горки с высотой (A’) и окружностью (A”) скважины, пробуренной в середине слайда. (B) (Вид снизу) Крышка, блокирующая скважину, герметизирована к слайду силиконовой смазкой. (C)(Вид сверху) Рассеченные яриоли развиваются в среде визуализации, которая покрыта газопроницаемой мембраной. Галогенуглеродное масло используется для стабилизации мембраны. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Чтобы проследить за ядерной миграцией и точно оценить траектории в ооците, необходимы маркеры как для ядерной оболочки, так и для плазматической мембраны. С этой целью были выбраны два трансгена, которые имеют высокое соотношение сигнал/шум и не исчезают в ходе живой визуализации. Для маркировки плазматической мембраны рекомендуется использовать P[ubi-PH-RFP], который кодирует домен гомологии Плекстрина (PH) фосфолипазы человека C ∂1 (PLC∂1), слитой с RFP. Этот домен PH связывается с фосфоинозитидом PI(4,5)P2, распределенным вдоль плазматической мембраны ооцита9. Для ядерной оболочки штамм P[PPT-un1]Fs(2)Ket-GFP, где GFP вставлен в ген, кодирующий Drosophila ß-importin, отображает однородный и интенсивный сигнал10. Молодых мух (1-2 дня) помещают в свежие флаконы, содержащие сухие дрожжи, за 24-48 ч до рассечения яичники.
Для этого анализа в реальном времени кусок алюминия толщиной 1 мм, который не является реактивным для образца, был разрезан на размеры слайда микроскопии. Он имеет отверстие диаметром 16 мм в центре затвора, которое было уравнёно до 0,85 мм. Этот контрбор имеет дополнительное отверстие диаметром 6 мм глубиной 150 мкм(рисунок 3А). Крышка наклеивается силиконовой смазкой (инертной для образца) на дно алюминиевой камеры(рисунок 3B). После помещения образцов в заполненную средой скважину над средой помещают мембрану, проницаемую для обменаO2/CO2 и окружают галогенуглеродным маслом(рис. 3С).
Для рассечения рекомендуется использовать щипцы из нержавеющей стали с размером наконечника 0,05 х 0,02 мм и иглы диаметром 0,20 мм для разделения яриолов(рис. 4В,С). Мигрирующие ядра визуалируются на вращающийся диск конфокального инвертированного микроскопа CSU-X1, оснащенного камерой. Многопозиционные изображения были получены с помощью покадровой съемки каждые 15 минут при 24 °C. Интервал в 15 минут позволяет выполнять многопозиционные захваты с ограниченным фотоотбеливанием флуоресцентных белков и фототоксичностью для образцов. Кроме того, более короткий интервал не даст гораздо более информативных данных для отслеживания ядерных траекторий. Фильмы обрабатываются и анализируются с помощью программного обеспечения Fiji11.
