Lignes transgéniques pour imager le cœur
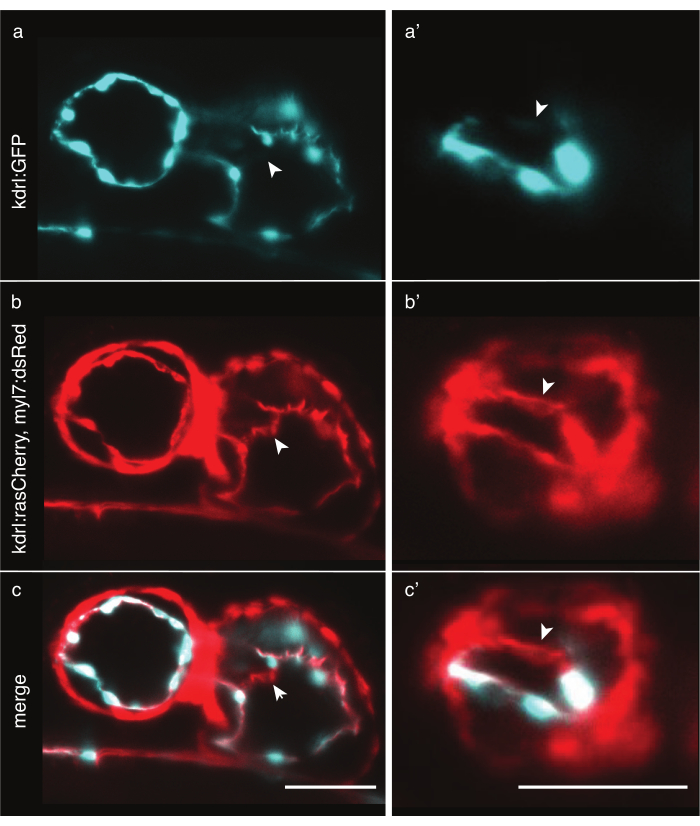
Figure 6 : Comparaison des lignées transgéniques de poisson-zèbre marqueur cytoplasmique et membranaire. Vue antérieure-ventrale de cœurs de poisson zèbre de 48 hpf imagés avec LSFM. Les flèches blanches indiquent des structures visibles uniquement avec une ligne transgénique marqueur membranaire. a) Signal Tg(kdrl:EGFP)32 en cyan dans le cœur et (a’) dans le ventricule. b) Signal Tg(kdrl:Hsa.HRAS-mCherry; myl7:dsRed)33 en rouge dans le cœur et (b’) dans le ventricule. ( c,c’) fusion des signaux Tg(kdrl:Hsa.HRAS-mCherry; myl7:dsRed) et Tg(kdrl:EGFP). Barre d’échelle 50 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
L’imagerie du cœur du poisson-zèbre nécessite un marquage précis des cellules cardiaques. Alors que l’épaisseur du myocarde est relativement constante dans toutes les cellules, les cellules endocardiques sont épaisses autour du noyau mais ont de minces protubérances membranaires, dans certaines régions plus minces que 2 μm. Les lignées transgéniques cytoplasmiques telles que Tg(kdrl:EGFP)32 marquent efficacement les régions autour des noyaux endocardiques, mais plus loin, le cytoplasme mince pourrait ne pas émettre suffisamment de photons pour être détecté avec des temps d’exposition aussi courts, ce qui entraînerait des trous artificiels dans les données (Figure 6a). En revanche, les lignées transgéniques marqueurs membranaires telles que Tg(kdrl:Hsa.HRAS-mCherry)33 peuvent étiqueter efficacement l’endocarde et révéler plus de détails (Figure 6b,c). Pour chaque expérience, choisissez soigneusement la lignée transgénique la plus appropriée.
Immobilisation du poisson-zèbre
Le choix de la technique d’immobilisation dépend de la durée de l’expérience et de l’âge du poisson à imager. La tricaïne a été couramment utilisée pour l’immobilisation du poisson-zèbre, principalement en raison de sa facilité d’utilisation. En effet, le simple fait d’ajouter 130 mg/L de tricaïne aux milieux piscicoles permet d’obtenir leur anesthésie en 10 min. Comme il peut entraîner des défauts de développement et affecter la physiologie cardiaque20,22, nous recommandons d’utiliser la tricaïne uniquement pour de courtes expériences (moins de 30 min). Pour une imagerie plus longue, les injections d’ARNm α-bungarotoxine au stade à une ou deux cellules paralysent les poissons jusqu’à 3 jours après la fécondation (dpf) sans affecter le développement cardiovasculaire ou la physiologie22.
Choisir les bons tubes FEP
Les tubes FEP sont disponibles en différents diamètres et épaisseurs. Pour imager 0-5 dpf poisson, 0,8 mm est un bon diamètre intérieur; choisissez soit des tubes à paroi épaisse de 0,8 x 1,6 mm, soit des tubes à paroi mince de 0,8 x 1,2 mm. Nous recommandons les tubes à paroi mince; cependant, les parois plus épaisses offrent une stabilité et une rigidité accrues, ce qui peut être important si la chambre d’échantillonnage a un fluide fluide qui pourrait perturber et déplacer un tube mince. Pour les échantillons plus grands, 1,6 x 2,4 mm et 2 x 3 mm peuvent être utilisés.
Échanges de température et de gaz
Un aspect essentiel du bien-être de l’embryon de poisson-zèbre est la température. Idéalement, maintenez le poisson à 28,5 °C pendant l’imagerie, car la température de l’environnement affecte le développement et la fréquence cardiaque34.
D’après notre expérience, l’échange d’oxygène à travers le bouchon d’agarose à 2% ne maintient qu’une fréquence cardiaque stable jusqu’à 3-4 dpf. Par conséquent, la coupe de trous dans le tube assure la diffusion de l’oxygène. Il peut également être nécessaire pour l’administration de médicaments à l’échantillon si vous le souhaitez.
Suspension du rythme cardiaque.
Les vitesses d’acquisition rapides des microscopes à feuille de lumière équipés de manière appropriée permettent d’enregistrer le cœur battant in vivo. Cependant, pour acquérir une pile z non perturbée, on peut ralentir ou arrêter le cœur. Cependant, l’arrêt du cœur entraîne une relaxation du muscle cardiaque et peut entraîner l’effondrement du cœur6. La suspension du rythme cardiaque peut être effectuée en utilisant des morpholinos, des basses températures, un inhibiteur de la contraction musculaire ou de l’optogénétique. Ces méthodes ont chacune leurs inconvénients et doivent être soigneusement évaluées pour chaque expérience.
L’injection de 4 ng de morpholino cardiaque silencieux (sih) au stade d’une cellule peut arrêter le rythme cardiaque en ciblant le gène tnnt2a crucial pour la formation de sarcomères35. Les poissons-zèbres sih n’ont pas de battement de cœur et ne survivent que jusqu’à 7 dpf, lorsque les embryons commencent à dépendre du sang circulant pour l’oxygénation. Comme la morphogenèse cardiaque est entraînée par des forces génétiques et biomécaniques36, ces poissons présentent des malformations cardiaques autour de 3 dpf.
Comme le flux de Ca2+ est sensible à la température, la température influence la fréquence cardiaque chez les poissons-zèbres embryonnaires21. Par conséquent, l’abaissement de la température dans la chambre d’imagerie ralentit le rythme cardiaque. L’arrêt du rythme cardiaque nécessite des températures inférieures à 15 °C. Comme les poissons-zèbres sont généralement maintenus à 28,5 ° C, ces températures basses ne peuvent être maintenues que pendant de brèves périodes (moins de 10 min).
Des médicaments tels que les inhibiteurs chimiques des contractions musculaires, le 2,3-Bu-tanedione 2-monoxime (BDM), peuvent être ajoutés au milieu du poisson-zèbre (50 nM37,38) pour suspendre temporairement le rythme cardiaque. BDM est pratique à utiliser car il arrête la contraction cardiaque en moins de 15 minutes et peut être lavé pour restaurer la fonction cardiaque. Cependant, comme le BDM modifie le potentiel d’action cardiaque, il doit être utilisé avec prudence37.
Enfin, le cœur des poissons-zèbres transgéniques exprimant des canaux ioniques à faible luminosité ou des pompes telles que la channelrhodopsine ou l’halorhodopsine dans leur myocarde peut être manipulé et arrêté en éclairant le stimulateur cardiaque au niveau du tractus d’entrée avec de la lumière39,7,40,41,9.
Perspective
Les outils et solutions optimisés présentés pour étudier le cœur du poisson-zèbre in vivo permettent une imagerie douce et à long terme de la dynamique cardiaque ultrarapide. L’intégration de l’échantillon peut être adaptée à différentes modalités d’imagerie, telles que la microscopie confocale, la microscopie à deux photons ou la tomographie par projection optique (OPT). La microscopie à feuille de lumière, cependant, est probablement la technique préférée qui offre une section optique à une vitesse suffisante pour capturer la dynamique du cœur. Bien que ce protocole se concentre sur l’imagerie cardiaque embryonnaire du poisson-zèbre, nous pensons qu’il pourrait également être appliqué à divers autres échantillons et expériences. Il sera intéressant de voir à l’avenir si des techniques d’intégration et d’imagerie similaires peuvent également être utilisées à des stades ultérieurs du développement, lorsque le cœur est plus caché et la larve moins translucide.
