All experiments were approved by the Institutional Animal Care and Use Committee at Cincinnati Children's Hospital Medical Center.
1. Preparation of D2O
NOTE: To avoid experimental variation, prepare sufficient solution/drinking water for all mice for the duration of the experiment.
- For intraperitoneal injection: generate 0.9% w/v saline D2O by dissolving 9 g of NaCl per liter of D2O. Filter through a non-pyrogenic 0.2 µm filter to sterilize.
- For drinking D2O-water: generate 8% v/v D2O-enriched water to be used as drinking water by mixing 80 mL of D2O per 920 mL of regular drinking water. Regular drinking water can be obtained from the mouse facility. Filter through a non-pyrogenic 0.2 µm filter to sterilize.
2. Modulation of BAT activity by temperature acclimation
- Separate the mice so there are two mice per cage 2 weeks prior to the start of temperature acclimation. Change the water supply to water bottles for the mice to adapt to and maintain all the cages at 22 °C.
- Prepare the mouse environmental chambers 1 week before starting the temperature acclimation by setting the appropriate temperatures: 30 °C for thermoneutrality, 22 °C for room temperature, and 18 °C for cold exposure.
- At the start of temperature acclimation, replace the cages with new ones with no environmental enrichment (to avoid nests). Move the cages to their respective temperatures.
NOTE: Cages assigned to thermoneutrality will stay at 30 °C for 4 weeks. Cages assigned to room temperature will stay at 22 °C for 4 weeks. Cages assigned to cold exposure will become progressively colder on a weekly schedule: 18 °C for the first week, 14 °C for the second week, 10 °C for the third week, and 6 °C for the fourth week. - Change soiled cages weekly for all conditions. Additionally, replace food and water bottles with new ones preadapted at the appropriate temperature for at least 24 h.
NOTE: Be mindful about the food amounts added every week, especially for mice in the cold, since they will consume a significantly greater amount of food compared to normal conditions. Provide food ad libitum.
3. Administration of D2O
- Inject each animal with 0.9% w/v saline-D2O at 35 µL/g of body weight, 12 h-3 days before tissue collection, via intraperitoneal injection using a 1 mL syringe and 26 G needle.
NOTE: Please see the discussion section for more information on selecting an appropriate labelling time. - Change the water bottles to bottles containing 8% v/v D2O-enriched drinking water.
4. Plasma and tissue collection, processing, and storage
- At the end of the experiment, sacrifice the mice using approved methodologies (e.g., carbon dioxide overdose followed by cervical dislocation).
- Use heat/cool pads or other methods to avoid sudden temperature changes before euthanasia that may affect the results. Euthanize the mice following approved methodologies. Proceed immediately to blood and tissue collection.
- Collect blood through cardiac puncture using a 26 G needle and store in an ethylenediaminetetraacetic acid blood collection tube. Keep the blood on ice until further processing.
- Cut open the skin along the middle line of the back of the mouse from the lower area of the thoracic cavity up to the upper area of the neck, while pulling the skin up to avoid affecting tissues below the skin. The interscapular BAT is located between the shoulder blades under a thin layer of white adipose tissue, and it is composed of two pyramidal shaped lobes.
- Clean the tissue by washing in ice cold phosphate buffered saline (PBS). Dab on a paper towel to eliminate excess liquid, weigh on an analytical scale, and collect in a microtube.
- Immediately flash freeze the tissue using liquid nitrogen. Other BAT depots can also be collected.
- Centrifuge the blood samples at 10,000 x g for 10 min at 4 °C. After centrifugation, carefully collect the plasma without disturbing the red blood cell pellet. Transfer it to a new ice-cold microtube and flash freeze in liquid nitrogen.
- Store brown fat and plasma samples at -80 °C until use.
5. Lipid extraction from adipose tissue
- Before beginning the extraction
- Prepare a 1 mM solution of hexadecenoic-d31 acid in methanol in a glass vial. This will serve as the internal fatty acid standard.
- Pre-cool the required amount of chloroform (CHCl3) and methanol (CH3OH) in a -80 °C freezer, or on dry ice.
NOTE: The antioxidant di-tert-butyl-4-methylphenol (BHT) may be added to the CHCl3 at a concentration of 0.01% w/v (2.5 mg/25 mL) to prevent oxidation of the double bonds in unsaturated fatty acids. - Pre-label microcentrifuge tubes for each tissue sample and an extra tube to be used for a blank extraction.
CAUTION: CH3OH and CHCl3 are highly volatile and toxic if inhaled. Use only in fume hoods.
NOTE: Different brands of microcentrifuge tubes have differing levels of background palmitate and capabilities in terms of preventing solvent leakage. We recommend that a variety of tubes be tested first, to ensure that solvent leakage is prevented and that the tubes have minimal levels of contaminating palmitate. Please see Yao et al.32 for further discussion.
- Take the samples out of the freezer and place on dry ice.
- Place the pre-labelled microcentrifuge tube on an analytical balance and tare the balance. Place tweezers and a scalpel/steel razor blade on dry ice for 10-20 s to cool.
- Use the tweezers to take the frozen tissue sample out of the tube and place on a plastic weigh boat. The weigh boat may be placed on a flat block of dry ice or another pre-cooled surface.
- Using the scalpel or steel razor blade, dissect a small portion of the tissue, equivalent to 5-15 mg in weight. Place in the microcentrifuge tube and record the exact weight. Repeat for each specimen. Samples can be stored in the freezer at this point or can be advanced to the steps below for lipid extraction.
NOTE: Ensure that the scalpel is properly cleaned with 70% ethanol between samples and a fresh weigh boat is used between each sample.
- Add 1 µL/mg of 10 mM hexadecenoic-d31(C16:0-d31) acid to each sample.
CAUTION: The following steps (5.4 to 5.8) should be carried out under a fume hood due to the inhalation risk of the solvents. - Add 250 µL of CH3OH, 250 µL of H2O, and 500 µL of CHCl3 to each sample with three 5 mm stainless steel beads. Place the tubes in a pre-cooled block of a grinding mill and mix the samples at a vibrational frequency of 25 Hz for 5 min, or use guidelines recommended by the manufacturer for tissue samples. Remove the beads using a magnet.
- Centrifuge the samples at 12,000 x g for 10 min at 4 °C.
NOTE: After centrifugation, a clear biphasic separation should be observed with the upper aqueous phase containing polar metabolites and the lower organic phase containing lipids and fatty acids. If no separation is seen, add 250 µL of H2O and repeat the vortex and centrifugation steps. - Using a micropipette, take a fixed volume from the bottom phase of each sample into correspondingly labelled microcentrifuge tubes.
- Add 500 µL of CHCl3 to the remaining sample and repeat steps 5.6-5.7.
- Place the samples under nitrogen gas or in a CHCl3-resistant refrigerated centrifugal vacuum at 4 °C until completely dry. Dried samples can be stored at -20 °C until ready for derivatization.
NOTE: The top layer of each sample can also be collected at this point and dried as in step 5.9 in order to analyze polar metabolites.
6. Preparation of fatty acid methyl esters (FAMEs) and GCMS analysis
- Acid-catalyzed esterification and transesterification to prepare FAMEs
CAUTION: The following steps should be carried out under a fume hood due to the inhalation risk of the solvents.- If the samples have been stored in freezer, dry under nitrogen for 5 min to ensure that no water is present.
- Using MS-grade solvents, pipette 98 mL of anhydrous CH3OH into a glass media bottle. Slowly add 2 mL of anhydrous sulfuric acid in the fume hood to make 2% H2SO4 in CH3OH. Mix by swirling the closed bottle.
- Add 500 µL of 2% H2SO4 in CH3OH solution to each sample and vortex briefly.
- Incubate the samples on a heat block at 50 °C for 2 h.
- Remove the samples from the heat block and add 100 µL of saturated NaCl solution and 500 µL of hexane to each sample.
- Vortex the samples vigorously at room temperature for 1 min. Leave the samples to sit for 1 min; two phases should be apparent after this.
- Collect the upper phase into a fresh microcentrifuge tube (see note in section 5.1.3 for appropriate selection of the microcentrifuge tube).
- To maximize the yield, repeat steps 6.1.5-6.1.7, collecting the second sample into the same labelled tubes.
- Dry the samples at room temperature under nitrogen gas.
- Resuspend the samples in 20 µL/mg of hexane, relative to the original tissue weight, and transfer immediately to glass GC vial with a glass insert.
NOTE: Work quickly while transferring samples to minimize evaporation.
- GCMS analysis
- To determine the abundance of FAME isotopologues, inject the samples on a single quadrupole gas chromatography mass spectrometer (GCMS).
NOTE: While many column types can be used to detect palmitate, the following temperature program was established for a GCMS column that has been developed for the separation of fatty acid cis/trans isomers, as detailed in the Table of Materials. This column has a length of 50 m with an 0.25 mm inner diameter. - Inject 1 µL of sample in a split/splitless inlet at an inlet temperature of 270 °C, using helium as the carrier gas, flowing at 1 mL/min. Use a splitless injection for low abundant fatty acids with a total flow of 19 mL/min, a septum purge of 3 mL/min, and a purge flow to split vent of 15 mL/min at 0.75 min. Use a split ratio of 10:1-40:1 for high abundant fatty acids such as palmitate and oleate.
- Use the following oven parameters: an initial temperature of 80 °C; increase by 20 °C/min increments to 170 °C; increase by 1 °C/min increments to 204 °C; increase by 20 °C/min increments to 250 °C; and then hold at 250 °C for 10 min.
- Use the following mass selective detector (MSD) parameters: an electron impact ionization mode of 70 eV and scan over the range of 50-400 m/z with a scan speed of 1,562 (u/s) and a frequency of 4.1 scans/s. Use a transfer line at 280 °C, an ion source at 230 °C, and a quadrupole at 150 °C.
NOTE: Other columns may be used, but the temperature program will vary. - Sample sequence: Randomize the sample order of injection and inject at least two or three hexane blanks at the beginning of the sequence, after every fifth sample within the sequence, and two or three at the end of the sequence.
- Use instrument specific software, or free open-access software such as Metabolite-Detector33, to integrate the ions in Table 1 for each fatty acid methyl ester.
NOTE: We have outlined ions that cover M1-M5 isotopologues in Table 1 that we have found covers the amount of deuterium incorporation with this labelling window. However, this may need to be expanded if de novo flux is significantly higher and/or a longer labelling time is used. - Use the abundance of each integrated ion to generate a mass isotopomer distribution, where the ion intensities can be converted to fractional abundance so that the sum of the mass isotopomer distribution equals one. Please see the example spreadsheet in the Supplementary File.
NOTE: In order to accurately determine deuterium incorporation, correction of the natural isotope abundance then has to be employed to allow for the presence of naturally occurring isotopes such as 13C, 15N, and 2H. This is performed by applying a correction matrix as outlined by Fernandez et al.34,35, and cannot be performed by subtracting the MID of an unlabelled measured metabolite from a labelled metabolite. In practice, we recommend the use of freely available software such as fluxfix36, polyMID37, or IsoCor38 to turn raw data into fractional MIDs, and we have provided the formula for isotope correction for the methyl ester products of FAS in Table 1. - To determine the quantity of palmitate present, utilize the following formula:
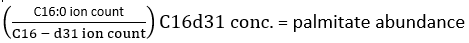
where ion count refers to the sum of all palmitate isotopologues integrated in Table 1 and C16:0-d31 refers to the internal standard hexadecenoic-d31. The internal standard forms a separate peak to that of the endogenous palmitate. The relative abundance of other fatty acids may also be determined, but fatty acid specific isotope internal standards (with mass shifts greater than that observed from D2O incorporation) or external standard curves may need to be employed for full quantitation. Use the blank extracted sample to determine the amount of background palmitate and subtract this from the final tissue value. - Calculate the molar enrichment (ME) of palmitate by the following equation:
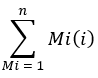
where Mi is the normalized, fractional abundance of a palmitate isotopologue, and n is the number of possible palmitate isotopologues. For example, the ME of a palmitate molecule with the following fractional distribution, M1 = 0.25, M2 = 0.08, M3 = 0.02, is: (0.025*1) + (0.08*2) + (0.02*3) = 0.245 (See Supplementary File).
- To determine the abundance of FAME isotopologues, inject the samples on a single quadrupole gas chromatography mass spectrometer (GCMS).
7. Deuterium acetone exchange of plasma samples to determine body water enrichment
- Reaction
- Prepare 10 standards of deuterium in water, ranging from 0-9% v/v.
- Prepare a 5% v/v acetone/acetonitrile solution allowing for 4 µL per sample, including the standards from step 5.1.1.
- In labelled, safe-lock microcentrifuge tubes, combine 10 µL of each plasma sample or standard, 4 µL of 10 M NaOH, and 4 µL of 5% acetone/acetonitrile. Perform this in triplicate for each sample.
- Mix the samples gently by pipetting. Allow the samples to incubate at room temperature overnight.
- Extraction
- After incubation, add 450-550 mg of Na2SO4 to each sample.
- In the fume hood, add 600 µL of CHCl3 to each tube and vigorously vortex for 15 s.
- Centrifuge the samples at 300 x g for 2 min.
- Under a fume hood, transfer triplicate, 80 µL aliquots of the supernatant from each sample into labelled, glass GCMS vials with glass inserts and cap tightly.
- GCMS analysis
- Separate samples on a column (30 m, 0.25 mm i.d, Agilent DB-35MS) and analyze on the attached mass spectrometer.
- Inject 1 µL of sample in a split/splitless inlet, with a split ratio of 40:1, a helium flow of 1 mL/min, and an inlet temperature of 270 °C.
- Use the following oven parameters: an initial temperature of 60 °C, increase by 20 °C/min increments to 100 °C, increase by 50 °C/min increments to 220 °C, and then hold at 220 °C for 1 min.
- Use the following mass selective detector (MSD) parameters: electron impact ionization mode at 70 eV with select ion monitoring of 58 and 59 m/z. Use a transfer line at 280 °C, an ion source at 230 °C, and a quadrupole at 150 °C.
NOTE: Other low-bleed, bonded, crosslinked, mid-polarity columns may be used, but temperature program will vary.
- Set the liquid sampler method to begin with a CHCl3 wash and then a randomized sequence of the samples, including additional CHCl3 wash steps every six samples.
NOTE: If acetone is used as a needle wash for the autosampler, replace with CHCl3 or hexane and inject multiple blank CHCl3 samples until any contaminating acetone peak is no longer evident in the chromatogram. - Using this method, an acetone peak elutes at around 1.25 min. Integrate the data and calculate the fractional abundance of acetone enrichment following steps 6.2.6-6.2.7, with an adjustment to analyze peaks containing an m/z ratio of 58-60 as indicated in Table 1.
- Use the standards to generate a standard curve to determine the percentage of body water enrichment (p) of the test samples, based on the fractional enrichment of acetone.
- Separate samples on a column (30 m, 0.25 mm i.d, Agilent DB-35MS) and analyze on the attached mass spectrometer.
8. In vivo de novo lipogenesis calculations
- Calculate the fraction of newly synthesized fatty acids (FNS) that are direct products of FAS (i.e., palmitate, odd chain fatty acids, and mmBCFAs) in each specimen with the following equation:
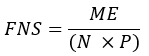
where ME is the average molar enrichment of a palmitate molecule (step 6.2.9), p is the deuterium enrichment in water from the corresponding plasma sample (step 7.3.4), and N is the number of exchangeable hydrogen atoms on palmitate where a deuterium can be incorporated. - Determine N using the equation below which was established by Lee et al.31:
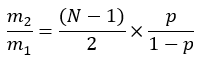
- Determine the molar amount of newly synthesized fatty acids (MNS) by:
MNS = FNS x total fatty acid amount (nmol/mg tissue).
NOTE: For example, if obtaining a palmitate molar enrichment (ME) of 0.245, a deuterium enrichment in body water (p) of 0.045, and a calculated N number of 22, the fractional synthesis of palmitate is 0.247. If the amount of palmitate present in the tissue is 2 mmol/mg, then the mmol of newly synthesized palmitate is 0.494 mmol/mg (see Supplementary File).
Based on the D2O dosing described in step 1, we typically find that body water is enriched in the range of 2.5% to 6%, and that a baseline level of deuterium enrichment in body water is rapidly achieved in 1 h and maintained for the duration of the study via 8% enriched drinking water (Figure 1). Continuous steady state body water enrichment is an assumption of the calculations used in step 6, and so we recommend experimental validation of the kinetics of body water enrichment in new experimental models.
We have found that the amount of de novo synthesized fatty acids is increased at room temperature, relative to those at thermoneutrality in BAT (Figure 2). The mass isotopologue distribution of palmitate in BAT from mice at thermoneutrality and room temperature after 3 days of D2O administration shows a higher M1 and M2 deuterium enrichment found at room temperature (Figure 2A). This colder temperature enrichment does not only happen in palmitate, but also in a broad range of fatty acids in BAT (Figure 2B). Total palmitate abundance is also increased in BAT of mice at room temperature, and combining the fractional synthesis rate with the total palmitate levels, we found that total palmitate synthesis was increased in mice at room temperature (Figure 2C,D). Notably, plasma total fatty acid enrichment does not follow the same trend as BAT, but instead, fatty acid enrichment is increased with thermoneutrality (Figure 2E). Deconvoluting potential uptake from endogenous synthesis is not possible with a D2O single time-point approach, as described here, but these opposing trends indicate that the fatty acid enrichment pattern in BAT is not being driven by fatty acid uptake.
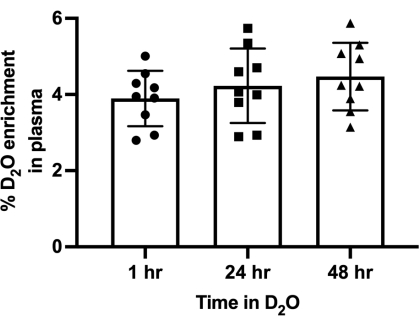
Figure 1: Percentage of D2O enrichment in plasma of mice over multiple time points, following injection with 0.035 ml/g body weight 0.9% NaCl D2O and replacement of drinking water with 8% D2O enriched water. Bar graphs represent mean ± SD. n = 9. Please click here to view a larger version of this figure.
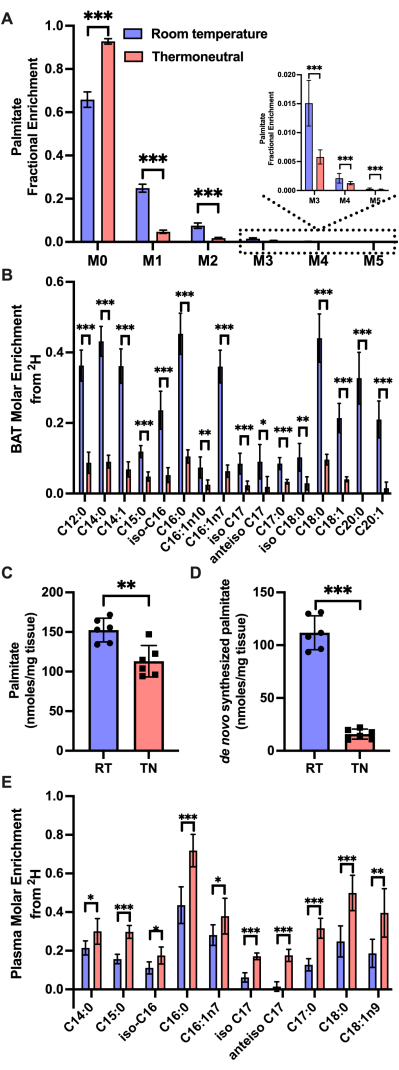
Figure 2: Representative results in mice brown adipose tissue after 3 days of D2O administration. (A) Mass isotopologue distribution of palmitate in BAT after 3 days of D2O administration at room temperature (RT) and thermoneutrality (TN). (B) BAT molar enrichment of a range of fatty acids after 3 days of D2O administration at room temperature and thermoneutrality. (C) Total abundance and the (D) de novo synthesized palmitate in brown adipose tissue after 3 days of D2O administration at room temperature and thermoneutrality. (E) Plasma molar enrichment of a range of fatty acids after 3 days of D2O administration at room temperature and thermoneutrality. Bar graphs represent mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001. n = 6. Statistical analysis was determined by two-tailed Students t-test. Please click here to view a larger version of this figure.
| Compound | Ions | Estimated elution time (minutes) | Formula for isotope correction |
| C16:0 | 270-275 | 20 | C17H34O2 |
| C14:0 | 242-247 | 16.5 | C15H30O2 |
| C15:0 | 256-261 | 18 | C16H32O2 |
| Iso-C16:0 | 270-275 | 18.9 | C17H34O2 |
| Iso-C17:0 | 284-289 | 21 | C18H36O2 |
| Anteiso-C17:0 | 284-289 | 21.5 | C18H36O2 |
| C16 D31 | 301 | 19.2 | ~ |
| Acetone | 58-59 | 1.5 | C3H6O |
Table 1: GCMS compound fragment ions to integrate. This table covers a range of fatty acids produced by mammalian fatty acid synthase, but C16:0 is the primary product. All retention times are for the GCMS method detailed in step 6 except acetone, which is detailed in step 7.
Supplementary File: Example spreadsheet calculation. Please click here to download this File.
