HLMEC isolation
The main problem during the isolation of HLMVECs is the presence of contaminating cells since the microscopic capillaries cannot be easily separated from the stromal tissue. Therefore, achieving the highest possible purity at the earliest stages of the isolation process is crucial in order to reduce the culture passages and, thus, the cell aging. Likewise, an optimal isolation protocol should give the highest possible yield of pure HLMVECs. To achieve these goals, a new procedure was set up based on previously described protocols10,11,12,14,15.
Although the lung parenchyma is highly vascularized, thus representing an excellent source of HLMVECs, it is largely populated by smooth muscle cells (SMCs), collagen fibers, and fibroblasts. These cells, especially SMCs, can adapt to a variety of culture media and, in almost all cases, replicate in vitro much faster than other cell populations, thus meaning they take over the mixed cell culture. Therefore, our first concern was to calibrate the enzymatic digestion of the parenchyma in order to reduce the carryover of unwanted cells. Since type 2 collagenase rapidly detaches groups of endothelial cells from the rest of the capillary, we reasoned that the digestion time of the lung sample could be crucial for minimizing cell contamination. Therefore, as a first step, the time of exposure to collagenase of the excised lung fragment was reduced to a maximum of 10 min instead of the suggested 20 min15 (Figure 2A). After this treatment, it was observed that part of the cell aggregates, including the SMCs and fibroblasts, were still bound to long fibers (Figure 2B), while SMCs represented the majority of non-blood cell singlets. Small, compact clusters of cells, with diameters generally not exceeding 10-20 µm, were also observed without elements attributable to fragments of stromal tissue (i.e., large fibers) (Figure 2B). It was hypothesized that these small cell aggregates could be composed mainly of HLMVECs.
Following this hypothesis, serial filtration steps were carried out to maximize the HLMVEC recovery. For the first filtration, a metal strainer was used, and the sample was massaged with tweezers to facilitate the escape of HLMVECs from the capillaries (Figure 2C, right). The aim of this step was to remove the large clusters of smooth muscle cells and fibroblasts still bound to the extracellular matrix. The outflow was then filtered using a sequence of 70 µm and 40 µm mesh size strainers to remove the residual large, although not eye-visible, aggregates (Figure 2C, middle, left). To remove the cell singlets, the samples were centrifuged at 30 x g for 5 min, a speed that sediments cells and aggregates with a diameter >7-10 µm, thus leaving the smaller cells in suspension. These centrifugation parameters were chosen based on those reported by Miron et al. for pelleting circulating blood cells22. Notably, after this centrifugation, the supernatant was red-colored (Figure 2D), indicating the presence of many red blood cells (~5 µm in diameter). At this point, the supernatant was carefully removed and placed in a second tube. Both tubes containing the supernatant and the pellet were filled with PBS−− and centrifuged at 300 x g for 5 min. After centrifugation, the tube containing the supernatant showed a red pellet enriched in blood cells, suggesting that it also contained most of the explanted cell singlets (Figure 2E, right).
The obtained pellets were suspended with the endothelial cell growth medium and separately seeded in two wells of a 6-well plate, which were pre-coated with 1.5% gelatin from porcine skin (Figure 2E, middle). After 3 days, the cells were washed with PBS++, fed with fresh medium, and imaged. As shown on the left of Figure 2E, EC islands were more abundant in the well containing the cells from "tube 1" (top), while the well containing the cells from "tube 2" (bottom) was mostly populated by single and non-endothelial-like cells.
The same procedure was applied to the less challenging HPAEC isolation protocol that was reported in our previous work, in which the HPAEC separation involved exploiting their shorter adhesion time compared with that of contaminant cells8. Due to their large size, arteries can be easily cleaned from residual connective and fat tissues, thus eliminating a large proportion of non-ECs. However, a large proportion of fibroblasts and extracellular matrix fibers was still present in the samples at this stage, in addition to vascular SMCs8. The new procedure further increased the initial EC purity during the HPAEC explants (results not shown).
Purification and phenotypic and functional characterization of freshly isolated HLMVECs
At the end of the isolation protocol, the purity of the HLMVECs was analyzed by the flow cytometric detection of cell membrane CD31. To correctly identify the CD31+ cell compartment, a gate was set on a forward-scatter (FSC)/side-scatter (SSC) dot plot, and a morphologically homogeneous population of cells was identified. Such a population was represented on an FSC-Area/FSC-Height dot plot, and single cells were gated and analyzed for the surface expression of CD31 (hierarchical gating strategy, not shown). In the best-case scenario, ~70% pure HLMVEC single cells (Figure 3A,B) were obtained. Of note, the HLMVEC yield was significantly reduced when the sample was not processed immediately after surgery or when the donor was elderly or a smoker.
To obtain purer HLMVEC preparations, the CD31+ cells were sorted and seeded in 1.5% gelatin-coated culture plates. The sorted cells assumed the characteristic endothelial-like shape (Figure 3C), and as reported in Figure 3D, confocal microscopy showed that almost 100% of the FACS-purified cells stained positive for EC antigens, namely CD31 and the more specific von Willebrand factor (vWF)23. Moreover, when these cells were seeded in Matrigel, they generated tubular structures, as well as HUVECs (Figure 3E), thus confirming their endothelial phenotype.
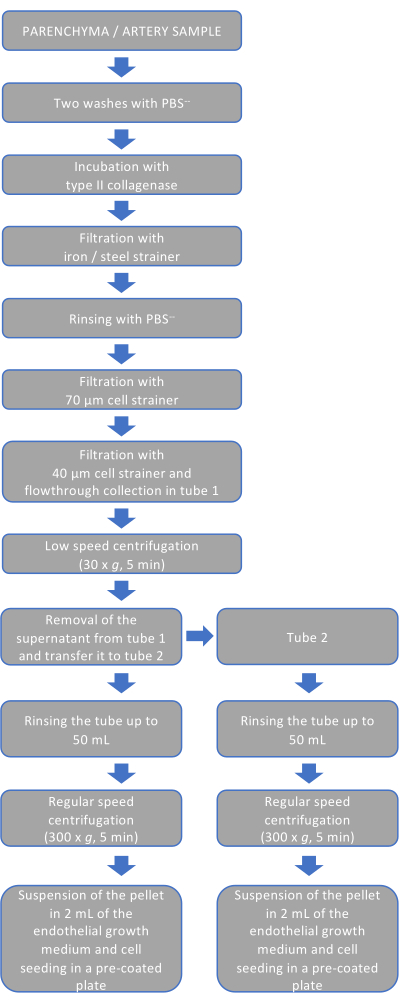
Figure 1: Schematic representation of the procedure. The flowchart shows the procedure for endothelial cell isolation and non-endothelial cell removal. Each step recapitulates the events described in steps 2-6 of the experimental protocol. Please click here to view a larger version of this figure.
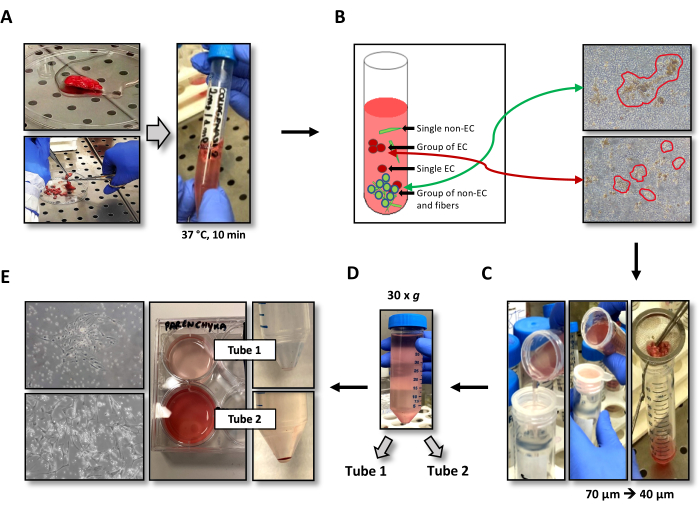
Figure 2: Illustration of the optimized EC isolation procedure. (A) Representative image of a lung parenchyma specimen diced in small fragments before collagenase digestion (left) and incubated in a 15 mL tube containing 5 mL of type 2 collagenase (right). (B) Schematic (left) and actual (right; magnification: 100x) representation of the cell populations obtained after collagenase digestion. These populations include singlets of non-ECs and ECs, groups of ECs, and groups of non-ECs bound to fibers, which are removed using strainers. (C) Serial filtrations using metal 70 µm and 40 µm mesh strainers. (D) Image of the 50 mL tube after low-speed centrifugation and before separating the supernatants from the pellets in "tube 1" and "tube 2". Cell clusters are contained in the pellet, while the single cells are mostly contained in the supernatant. (E) Image showing the different colors and compositions of the pellet and the isolated cells from "tube 1" (top) and "tube 2" (bottom); magnification: 100x. Abbreviation: EC = endothelial cells. Please click here to view a larger version of this figure.
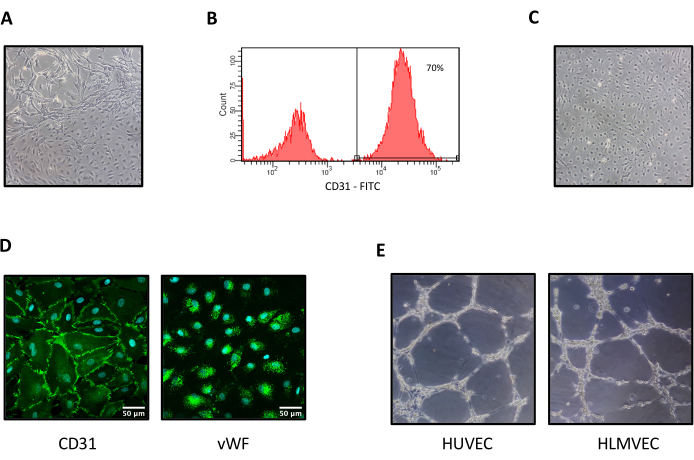
Figure 3: HLMVEC phenotypic and functional characterization. (A) A mixed cell population obtained from a lung parenchyma explant. (B) CD31 detection by flow cytometry on the surface of cells at the end of the first passage. The percentage purity (70%) refers to the used hierarchical gating strategy. (C) Pure HLMVECs after being live-sorted for the CD31 antigen. (D) Representative immunofluorescence confocal microscopic images of CD31 (left) and vWF (right) positive staining (scale bar: 50 µm). (E) Brightfield images were captured 6 h after seeding HUVECs or HLMVECs on Matrigel.Magnification (A, C, E): 100x. Abbreviations: HLMVECs = human lung microvascular endothelial cells. Please click here to view a larger version of this figure.
