The yeast Saccharomyces cerevisiae is a widely recognized model used to study mitochondrial biogenesis. Since yeast is an anaerobic, facultative organism, it is possible to extensively study the causes and consequences of introducing mutations that impair respiration. In addition, this organism possesses friendly genetic and biochemical tools to study mitochondrial pathways. However, one of the most powerful resources to explore the mechanisms of respiratory complex assembly and mitochondrial protein synthesis is the ability to transform mitochondria and modify the organelle's genome. Previously, it has been helpful to introduce in the mitochondrial DNA (mtDNA) point mutations or small deletions/insertions1,2,3,4,5, delete genes6,7, make gene rearrangements7,8, add epitopes to mitochondrial proteins9,10, relocate genes from the nucleus to the mitochondria11,12, and introduce reporter genes like BarStar13, GFP14,15, luciferase16, and the most widely used ARG8m17,18. Mitochondrial genome modifications have allowed us to dissect and identify mechanisms that otherwise would have been difficult to comprehend. For example, the ARG8m reporter gene inserted at the COX1 locus in the mitochondrial DNA was crucial to understanding that Mss51 has a dual role in Cox1 biogenesis. First, it is a translational activator of the COX1 mRNA, and second, it is an assembly chaperone for the newly made Cox1 protein7,19. This work presents a detailed method to transform S. cerevisiae mitochondria. Although the mitochondrial transformation protocol was published earlier16,20,21,22,23, a visual approach through a video is essential to thoroughly understand the different stages and details of the method. The method consists of various steps and is divided into four general stages:
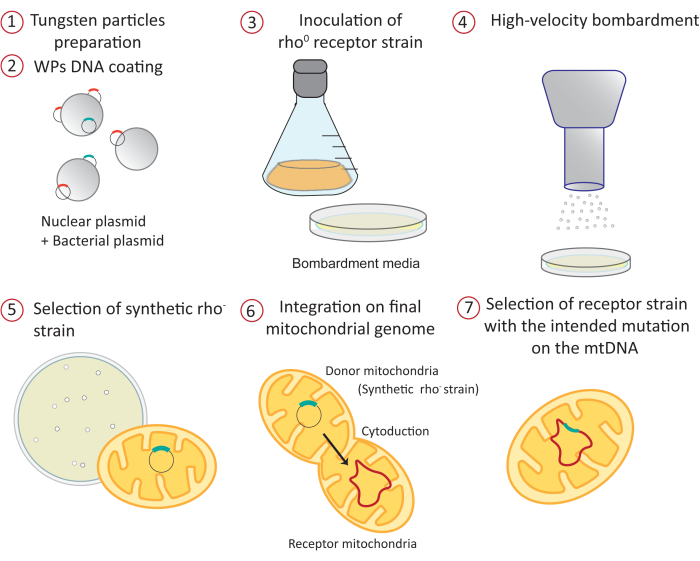
Figure 1: Overview of the mitochondrial transformation procedure by high-velocity microprojectile bombardment: 1) The tungsten particles are sterilized and prepared before DNA coating. 2) Two different plasmids are precipitated on the surface of the WPs. One is a bacterial plasmid containing the construct that will be directed to the mitochondrial matrix. The other is a nuclear yeast expression vector carrying an auxotrophy marker. 3) A receptor yeast strain lacking mitochondrial DNA (rho0) is grown on a fermentative carbon source like galactose or raffinose, which does not exert any glucose repression of mitochondrial gene expression34,35. The culture is spread on petri dishes containing the bombardment medium. 4) WPs coated with plasmids are shot to the receptor strain by high-velocity microprojectile bombardment. 5) Positive synthetic rho– colonies containing the mitochondrial plasmid are selected by mating to a tester strain. 6) The mitochondrial construct is integrated into the mitochondrial genome at the desired locus by mating the synthetic rho– strain (donor) with the acceptor strain by a process known as cytoduction. 7) The positive receptor, haploid strain carrying the mitochondrial construct is selected and purified in different media. Abbreviations: WPs = tungsten particles; mtDNA = mitochondrial DNA. Please click here to view a larger version of this figure.
Transformation of cells with the DNA construct intended to integrate into the mitochondrial genome (Figure 1, steps 1-4)
Although it is possible to directly transform a yeast containing a complete mitochondrial genome (rho+), the transformation is 10-20x more efficient if the cells lack mitochondrial DNA (rho0)24. Tungsten particles (WPs) are coated with two different plasmids that will be co-transformed. The first one is a yeast 2 µ expression vector carrying an auxotrophy marker, such as LEU2 or URA3 (e.g., YEp351 or YEp352, respectively). If the biolistic introduction of DNA into the yeast cells is successful, the transformants will grow on the auxotrophy medium (i.e., a medium lacking leucine or uracil). This plasmid is helpful in making the first selection of the cells that acquired the nuclear plasmid; otherwise, the number of resulting colonies would saturate the plate. The second plasmid is a bacterial plasmid (such as pBluescript or similar) containing the mitochondrial construction intended to be integrated into the mitochondrial genome. The construct must contain at least 100 nt of 5' and 3' flanking mitochondrial sequences for recombination with the mitochondrial region of interest. In our experience, larger flanking sequences are more likely to successfully recombine with the target locus in the mitochondrial genome.
A specific example is shown in Figure 225. In this example, the aim was to delete the region of the mitochondrial COX1 gene coding for the last 15 amino acids of the corresponding protein (Cox1ΔC15) (Figure 2A). The bacterial plasmid carrying the COX1ΔC15 mutation contained 395 nt and 990 nt of the gene's 5' and 3' untranslated regions, respectively. The plasmid was derived from pBluescript (pXPM61) and, together with the 2 µ plasmid YEp352, they were coprecipitated on the surface of WPs (Figure 2B). The WPs were then introduced by microprojectile bombardment into the selected recipient strain (Figure 2C). This strain, named NAB6926, is a MATa, rho0 strain bearing the kar1-1 and ade2 alleles (the importance of these characteristics is discussed below). The cells were plated on bombardment media lacking uracil to select those that acquired the 2 µ plasmid (Figure 2C). Cells were grown for 5-7 nights at 30 °C.
Selection of the cells that acquired the bacterial plasmid containing the mitochondrial construct and the nuclear 2 µ plasmid carrying the auxotrophy marker (Figure 1, step 5)
The positive colonies will maintain many copies of the bacterial plasmid in their mitochondria22. Since the transformed cells are originally rho0, no mitochondrial sequences support the translation of the mitochondrial construct present in the plasmid; hence, the transformed mitochondrial gene will not be expressed. It is necessary to mate the transformants with a tester strain to detect the yeast colonies that acquired the mitochondrial plasmid. The mitochondrial genome of the tester strain contains a nonfunctional mutated version of the gene of interest. After mating, the mitochondria will fuse, and the mitochondrial sequence included in the transformed plasmid will recombine with the mutated mitochondrial gene from the tester strain; consequently, the recovery of the WT gene will reconstitute function. The resulting diploid will have a detectable positive phenotype (usually the capacity to grow in respiratory medium or a medium lacking arginine). The positive cells carrying the bacterial plasmid in mitochondria are named "synthetic rho– cells". In the specific example of Figure 2D, synthetic rho– cells carrying the COX1ΔC15 construct (named XPM199) were replica-plated on a medium lacking uracil (this is the master plate from which positive colonies were purified). They were also replica-plated on a tester strain L4527 lawn, which contains the nonfunctional mutation cox1D369N. After mating for two nights, the mitochondrial genome of L45 and XPM199 recombined, resulting in a functional COX1 gene; therefore, the diploids recovered the ability to grow on respiratory media. From the master plate, we picked the positive colonies. They were streaked on plates lacking uracil, and the selective tests were repeated to obtain pure synthetic rho– cells (Figure 2E). It is important to note that the testing plate is only for the identification of synthetic rho– colonies, and mutants cannot be recovered from these plates.
Integration of the construct into the mitochondrial genome of the intended strain
This step is achieved by a process called cytoduction28,29 (Figure 1, step 6). In this approach, the synthetic rho– strain (donor strain) is mated to the intended acceptor strain of the opposite mating type. An essential requirement is that at least one of the two mating strains carries the kar1-1 mutation to impede nuclear fusion30. Hence, mating of the two cells produces a binucleated zygote (Figure 3). The mitochondrial network from the parental cells (donor and acceptor) fuse and the mitochondrial DNA molecules recombine. The binucleated zygotes are incubated/recovered to allow budding, which will produce haploid cells. These haploids carry the nuclear background of one of the parental cells. Similarly, haploids can carry the mitochondrial DNA from either the parental cell or the recombined mitochondrial DNA of interest. However, the kar1-1 mutation is not 100% effective, and some true diploids will be formed during the mating28,29. The synthetic rho– strain (donor, XPM199) carried the kar1-1 allele in the example. As a selective marker, it had the ade2 allele, making it an auxotroph for adenine (Figure 4). The acceptor strain was XPM10, a rho+ strain bearing the cox1Δ::ARG8m construct, where the reporter ARG8m replaced the COX1 codons in the mitochondrial genome7. The mixture of both cell cultures was added to a YPD plate as a drop to allow mating. After 3-5 h, the cells were observed under a light microscope to detect the formation of shmoos (Figure 3B), a cell shape associated with mating. After the incubation/recovery time, the binuclear zygotes were plated on a medium lacking adenine to prevent the growth of the donor cells.
Selection of the haploid strain carrying the mitochondrial genome of interest (Figure 1, step 7)
A mixture of donor, acceptor, and diploid cells is present after cytoduction. Therefore, after the incubation/recovery of the binucleated zygotes, cells must be grown in different selective media to identify and purify the haploids of interest. The selective media depends on the genotypes of the donor, acceptor strains, and the intended mitochondrial construction. However, in general, the reasoning for choosing the selective media is: i) after incubation/recovery, the cytoduction mix is grown on selective medium where the donor parental strain cannot grow. In the specific example in Figure 4A,B, the donor (synthetic rho– strain) carries the nonfunctional ade2 allele. Thus, the cytoduction mix must be incubated on a medium plate lacking adenine. This is the master plate. ii) Once the colonies grow, the master plate is replicated again on a medium lacking adenine (to generate a new master plate from where the positive colonies of interest will be purified), and next, on a medium where only diploids can grow. This is necessary to avoid further inadvertent purification of diploid colonies. In the case of the example from Figure 4C, only diploids can grow on media lacking leucine. iii) The master plate is also replicated on media where only those acceptor haploids that incorporated the mitochondrial DNA of interest will grow. In the example of Figure 4C, haploids grow on respiratory media containing ethanol/glycerol as a carbon source since the Cox1ΔC15 protein is functional. The resulting rho+ strain bearing the mtDNA of interest was named XPM20925. The strains used in Figure 2 and Figure 4 are listed in Table 1.
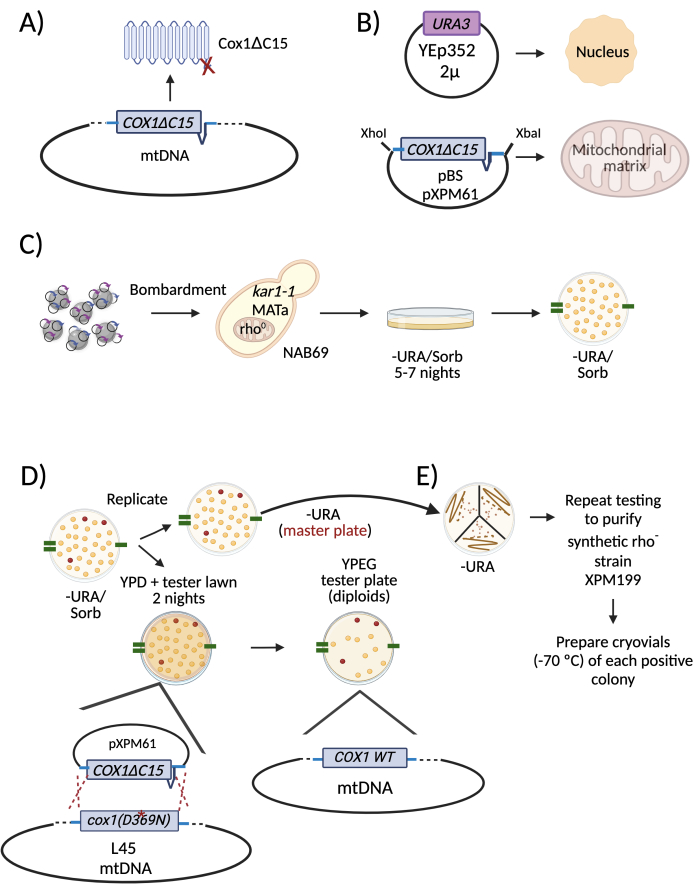
Figure 2: Diagram describing a specific example of mitochondrial transformation and selection of positive rho– cells. (A) The intended modification of the mitochondrial genome was a deletion of the region coding for the last 15 amino acids of the Cox1 subunit (Cox1ΔC15). (B) WPs were coated with two plasmids to transform yeast cells. One was the 2 µ plasmid Yep352 that was directed and expressed in the nucleus. The other was plasmid pXPM61, which contains the COX1ΔC15 allele plus 395 nt and 990 nt of the COX1 5' and 3'-UTRs, respectively. (C) The WPs were introduced into cells from the strain NAB69, which lacks mitochondrial DNA (rho0 strain). Transformant colonies obtained from the bombardment plates were replicated on a medium lacking uracil, the auxotrophic marker of the nuclear plasmid YEp352. (D) To select the positive rho– colonies, the -URA plate was replicated on a media lacking uracil. This was the master plate from which the positive colonies were picked and isolated. It was also replicated on plates with rich media (YPD) and a lawn of a tester strain. During mating, the synthetic rho– mitochondrial DNA was recombined with the mitochondrial DNA from the tester strain, L4527, which carries a mutation in the COX1 gene (D369N). The diploids that grew on respiratory media contained the transformed DNA. The corresponding colonies were picked from the master plate to restreak and purify. To help identify the positive colonies in the master plate throughout all replica plating, marks were made with a permanent marker on the edges of the plates (green lines). (E) The selected positive colonies were restreaked on a medium lacking uracil. This was the new master plate. As in D, two more rounds of testing were carried out to purify the synthetic rho– colonies. Abbreviations: WPs = tungsten particles; mtDNA = mitochondrial DNA; -URA/Sorb = lacking uracil/ containing Sorbitol; WT = wild type. Please click here to view a larger version of this figure.
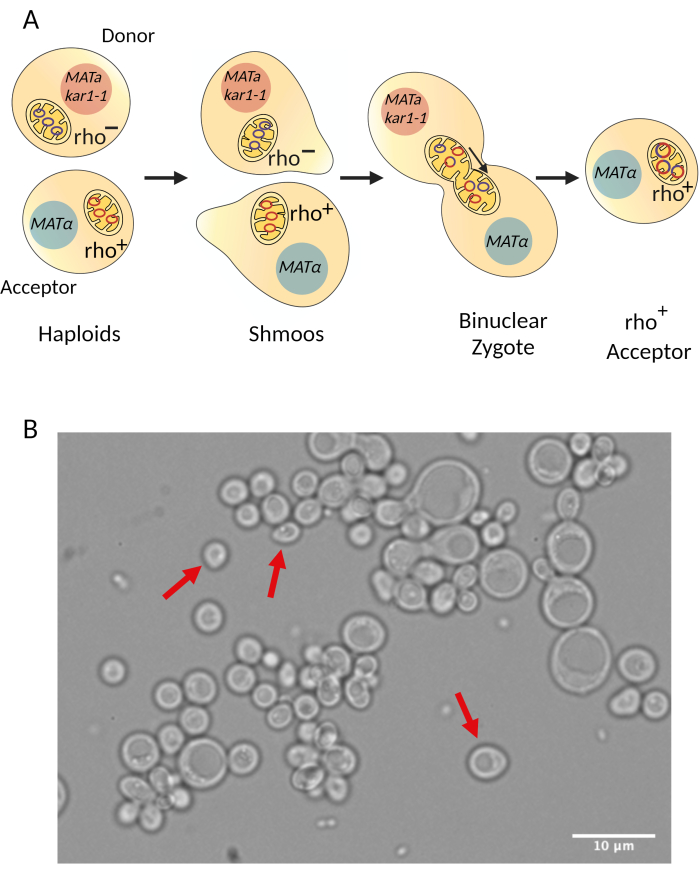
Figure 3: Diagram describing the cytoduction procedure. (A) General overview of how cells mate during cytoduction. During cytoduction, mitochondria from the donor and acceptor strains fuse so mitochondrial DNA from both strains recombine. Since nuclear fusion is reduced due to the kar1-1 mutation30, binuclear zygotes are formed. After some incubation/recovery time, the binuclear zygotes bud and haploid acceptors containing the intended mitochondrial mutation are selected. (B) Mating of the synthetic rho– strain (donor) and the acceptor strain through cytoduction allows the formation of shmoos (red arrows), a characteristic shape of mating yeast. The image was taken under a light microscope with a 100x objective. Scale bar = 10 µm. Please click here to view a larger version of this figure.
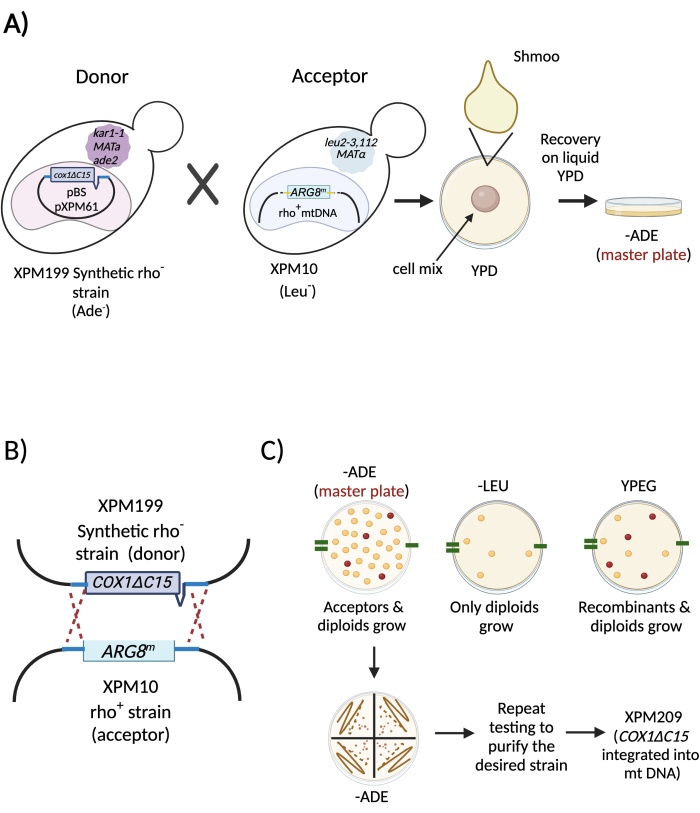
Figure 4: Diagram describing a specific example of cytoduction to integrate the mutation COX1ΔC15 in the mitochondrial genome. (A) Liquid cultures of the synthetic rho– strain (donor, XPM199) and the strain of interest (acceptor, XPM10) were mixed to mate. After shmoo formation, the mix was recovered in a liquid culture for 2-4 h. Next, the cytoduction mix was spread on a plate with a medium lacking adenine to prevent the growth of the donor cells. (B) Schematic representation of the recombination events of the mitochondrial DNA from the donor (XPM199) and the acceptor (XPM10) during cytoduction. (C) The master plate was replicated on different selective media to identify and purify those haploids that integrated the intended mitochondrial gene in the organelle's DNA (XPM209)25. Abbreviations: -ADE = lacking adenine; -LEU = lacking leucine. Please click here to view a larger version of this figure.
