At the heart of the adaptive immune system lies the ability of a T cell to recognize and respond to a specific antigen. When and where a T cell responds to its cognate antigen determines the balance of infection and autoimmunity, homeostasis and cancer, health and disease1. It follows that the study of T cells in a specific context of immunity should focus on the cells with specificity for a relevant antigen of interest. Among the technological advancements that have greatly enhanced the ability to characterize antigen-specific T cell populations are fluorescently labeled soluble multimers (usually tetramers) of major histocompatibility complex (MHC) class I or class II molecules complexed to antigenic peptide epitopes, better known as "peptide:MHC tetramers"2,3,4,5. By representing the natural ligands of T cell antigen receptors (TCRs), peptide:MHC class I and class II tetramers provide a means to directly identify antigen-specific CD8+ and CD4+ T cells, respectively, within the endogenous repertoire of T cells in the immune system without a requirement for a response to antigen stimulation in an assay. Tetramers represent a more elegant approach to the study of antigen-specific T cells than TCR transgenic T cell adoptive transfer models6, and have been increasingly used to identify foreign and self antigen-specific T cell populations in both experimental mouse models and human disease4,5.
While tetramers can readily identify high-frequency populations of T cells that have expanded in response to antigen stimulation, their use for naive, self antigen-specific, or memory T cells is limited by the very low frequencies of these populations7. Our group and others have developed and popularized tetramer-based magnetic enrichment strategies that increase the sensitivity of detection to enable studies of these cell populations in mouse lymphoid tissues8,9,10,11.
The emergence of tissue-resident T cells in the field has placed a heightened emphasis on developing new ways to investigate T cells in the nonlymphoid space. Like many other mucosal surfaces, T cells in the lungs encounter a range of self and foreign antigens derived from host epithelium, commensal and infectious microbes, and environmental entities, including allergens. Transcriptional analysis of T cells harvested from nonlymphoid tissue (NLT) demonstrates a memory-like phenotype that bears unique tissue-specific fate and function, often directed at trafficking and tissue homeostasis12. Moreover, tissue-resident memory T cells (Trms) tend to be more clonally restricted than those in circulation13. Determining how and why antigens drive T cell residence in NLT is critical to understanding how the immune system protects against infection, maintains tissue homeostasis, and, at times, devolves into autoimmunity. However, there appears to be greater attrition amongst tissue-resident T cells from the lungs compared to other NLT14. Accordingly, the ability to identify and characterize endogenous T cells of the lung with a given antigen-specificity is limited by their inherent rarity.
By combining the use of magnetic bead-based cell enrichment techniques and peptide:MHC tetramer staining, we have succeeded in detecting expanded but rare self antigen-specific T cells in mouse lungs15,16. Here, we present a detailed description of a protocol that we have optimized to reliably isolate and characterize any rare antigen-specific T cell population present in mouse lungs (Figure 1). This protocol incorporates an in vivo antibody staining step to distinguish tissue-resident from vascular T cells17 followed by two different methods for lung tissue processing to accommodate resource availability. This is then followed by a general T cell magnetic enrichment step, tetramer staining, and analysis by flow cytometry. Cell viability and tetramer staining is further enhanced in this protocol by the addition of aminoguanidine, which blocks inducible nitric oxide synthase (iNOS)-mediated T cell activation-induced apoptosis18 and Dasatinib which limits TCR downregulation19. The steps highlighted in this protocol utilize common techniques and readily available reagents, making it accessible for nearly any researcher engaged in mouse T cell immunology and is highly adaptable for a variety of downstream analyses. Although naive T cells are not likely to be found in the lungs, we believe this protocol will be particularly helpful for the study of self antigen-specific T cells and Trms in the lungs.
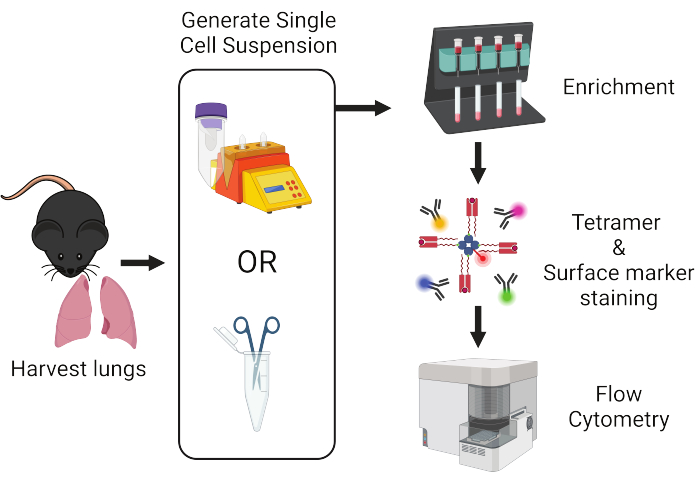
Figure 1: Overview of protocol workflow. Lungs are harvested from mice and dissociated into single cells. Samples are subsequently enriched for T cells prior to staining with peptide:MHC tetramers and fluorescently-labeled antibodies for flow cytometric analysis. Please click here to view a larger version of this figure.
