To investigate the neurobiology of learning and memory, invasive techniques are required, which are not generally feasible in humans. Thus, for over a century, behavioural protocols have been designed for laboratory animals to model various forms of human memory. The design and choice of both task and apparatus are central to the success of effective models of human memory. Numerous paradigms have been developed with diverse complexity, ranging from simple classical and instrumental conditioning protocols1,2,3 to mazes such as the T-maze4, radial arm maze5, Barnes maze6, watermaze7, and the cheese-board maze8. Yet, while these tasks capture facets of associative learning and spatial navigation, they cannot be used unambiguously to study the memory representation of momentary events (i.e., episodic-like memory). And, although novel object recognition9 and permutations of this spontaneous memory task, such as object-place memory10, have provided valuable insights into recognition memory, they do not test explicit recall of events. To address this demand, the event arena was specifically developed, and its use has enabled research into long-term, paired-associate memory encoding and recall11,12,13 as well as the encoding and recall of discrete events happening in a familiar space14,15,16,17,18. The latter theme is the focus of this manuscript.
The event arena is a large, square, open-field area where events occur for rodents. The size of the arena can be scaled to accommodate either rats or mice, and rodents are encouraged to enter and explore. A typical example of an event that takes place within the arena is the finding and retrieval of food from a sandwell at a specific location. The event arena is designed for such appetitive tasks, in which rats or mice are trained to search for, find, and dig up food. It capitalizes on their natural tendency to carry food back to a dark environment, which in this case is located adjacent to the arena, where they then eat it. After minimal training to dig for food, rodents take to this task naturally and perform well in the encoding trial, and in the recall choice trial, which follows the encoding trials after a short, 30-min delay. In a choice trial, several sandwells (i.e., locations for digging) are available, but only one is rewarded.
Different tasks can be performed within the event arena (e.g., spatial memory, episodic-like memory, and paired-associate learning). Given the interest in developing effective models of episodic-like memory, the following protocol was developed, in which the location where food can be found is altered daily. In this task, rodents are required to remember where the event of digging for, and successfully retrieving, a food reward happened most recently within the event arena. The protocol outlined below entails an encoding trial in which rats search for a sandwell in a new place each day followed, after a delay, by a recall choice trial, where the recently encoded sandwell location is rewarded, while the other, alternative sandwells in different locations do not contain accessible food. Remembering where the food was on a previous day is not helpful: the correct location has to be encoded and remembered, at least for a while, each day. Accordingly, we have introduced the term 'everyday memory' to capture the form of memory modeled in this task, which we, as humans, use on a daily basis. A human example of everyday memory is remembering where one has parked one's car at the shopping mall (Figure 1A) or has put one's glasses down around the house. In this protocol, all intra- and extra-arena cues are all stable, just as they are in the settings of our everyday lives (i.e., homes, offices, car parks, etc.). Thus, rodents must remember where something happened most recently within a familiar environment (Figure 1B). The task is analogous to, but an improvement on, the delayed-matching-to-place (DMP) task in the watermaze19. Being an appetitive task, it exploits rodents' natural behavior to forage for food20, instead of their desire to escape from the water. However, as in the watermaze7, there are no local cues differentiating correct from incorrect locations; animals must use recall rather than recognition to locate the correct sandwell location after varying memory delay durations.
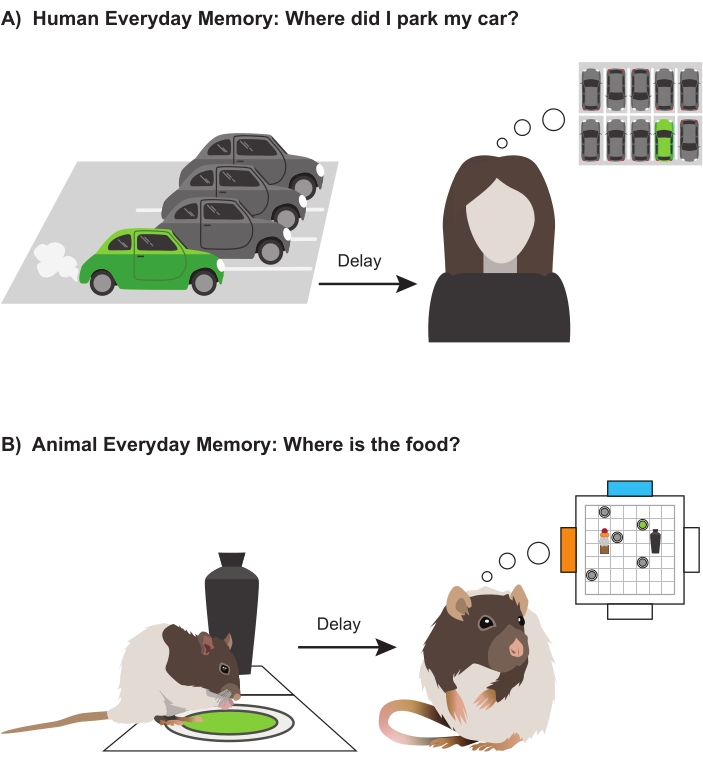
Figure 1: Everyday memory. (A) Human everyday memory. Schematic showing a green car parked in a car park. After a delay, the driver attempts to remember exactly where she parked her car. (B) Animal everyday memory. Schematic showing a rat digging and retrieving a pellet from a sandwell at a location within the event arena. After a delay, the rat is given a choice trial with multiple incorrect sandwells (gray) and one correct sandwell (green). Please click here to view a larger version of this figure.
The event arena has already been successfully utilized in investigations of 'everyday memory'. These are memories that are automatically encoded each day, retained in long-term memory, but often forgotten after relatively short time periods. Bast et al.14 showed monotonic delay-dependent event memory, which varies from excellent memory after short intervals to chance level after 24 h. The retention of memory can, however, be successfully enhanced by post-encoding novelty or, with multiple encoding trials, with extended trial spacing15,17.
The event arena is versatile and relatively non-stressful; no aversive stimuli are used. The size of the arena, and the tasks it accommodates, can be adapted for both rats14,15 and mice16. Also, as a land-based task, it is amenable to physiological recording and calcium imaging studies, unlike the watermaze21. Moreover, in accordance with the principles of the 3Rs (reduction, refinement, replacement), studies employing the event arena require fewer animals to obtain statistical power, as within-subject experimental designs are feasible (in which each animal serves as its own control for pharmacological interventions, optogenetic stimulations, etc.) and no aversive stimulation is required for motivation. Although initial training demands more time and occurs over more sessions than in, for example, novelty recognition tasks, once animals achieve a stable, asymptotic level of task performance, manipulations such as drug, vehicle-control, or optogenetic stimulation may be interspersed with a relatively small number of additional training sessions17. In addition, distinct facets of representation come under direct experimental control in the event arena, such as the nature of the spatial representation employed when solving the task.
The issue of representation concerns the mental framework employed by rats when remembering where recent events happen18. Do they remember where the food is located, or do they only remember how to get to the food? Rats can use allocentric (map-like) or egocentric (body-centered) spatial representations to solve an appetitive task within the arena18. However, to control and identify the spatial strategy employed by each experimental subject when performing the task, there are distinct training protocols that are able to selectively promote the use of only one spatial representation. Usually, an egocentric-based representation is employed when rats take their food reward back to the same location from which they started the day's trial, which allows several opportunities to remember the reward location during runs back and forth. This spatial strategy can be employed regardless of whether the start location is changed from day to day or kept constant. In contrast, an allocentric representation is favored when rats are required to carry food reward to a fixed home-base location at the side of the arena, which is different from the changing starting locations. There are numerous advantages of allocentric representations with respect to the brain's storage capacity.
In this paper, we have outlined the home-base protocol, which encourages the employment of only an allocentric representation. We have provided representative results for this task, which clearly illustrate the advantages of using this rodent model of 'everyday memory' in the investigation of learning and memory and highlights how allocentric representations of episodic-like spatial memory can be promoted.
The methods described in this paper have been approved by the University of Edinburgh Ethical Review Committee; they are compliant with the UK Animals (Scientific Procedures) Act 1986 and the European Communities Council Directive of 24 November 1986 (86/609/EEC) legislation governing the maintenance of laboratory animals and their use in scientific experiments.
NOTE: The experimental subject of the protocol outlined below is Lister-hooded rats, but it can be adapted for other rodent strains.
1. Animal handling, housing, and food control
- Allow 1 week for Lister-hooded male rats to settle after arrival. During this time, handle them daily by gently stroking and tickling them in their cages. Once they are settled, start picking them up for approximately 5 min each day.
- Record each rat's weight upon arrival and every 2-3 days per week. Tailor their food intake, so that each rat's weight is gradually reduced to approximately 85%-90% of their free-feeding body weight; this can be estimated using an established growth curve for free-feeding rats. Maintain the rats within this food-restricted weight range throughout the experiment.
- House the rats in a 12 h (light on)/12 h (light off) light cycle and conduct all experiments during the light phase (7 am-7 pm).
2. Setting up the apparatus
- Experimental rooms, control rooms, and event arena
- Experimental and control rooms are either one room separated into two parts or two adjacent rooms separated by a curtain or door and are required for this experiment.
NOTE: This separation will prevent the experimenters from influencing or disrupting the animals while they perform and learn this complex behavioral task. - Dedicate one room to the event arena, the static environmental cues, and the experimental procedures (i.e., the experimental room), and use the other room to record the rats' performance by the experimenters (i.e., the control room).
NOTE: An event arena is a square-shaped, open-field area where event-place associations can be studied (Figure 2A).The name 'event arena' derives from the fact that this apparatus is an arena (i.e., an open but constrained space) in which 'events' happen (e.g., digging up buried food pellets in sandwells; Figure 2B)17,18. To prevent overwork and injury (i.e., back strain) to the experimenter, the arena is elevated above the floor (~1 m). - Using transparent Plexiglass, build a square (160 cm x 160 cm) arena. The floor of the arena comprises a 7 x 7 grid of 49 movable white Plexiglass tiles (20 cm x 20 cm; Figure 2A,B). Modify five additional tiles with a central hole (6 cm diameter): these will hold the sandwells within the arena. The five locations of these modified tiles are arranged in the configuration outlined in each session's sandwell map.
NOTE: The configuration of the five sandwell locations changes with each new session (see step 4.2). - Place the event arena in the designated experimental room. Set up the intra-arena cues: position two distinctive landmarks with distinct tactile surfaces (e.g., a glued stack of golf balls (30 cm (h) x 11 cm (w) x 11 cm (l)) and a black water bottle (22 cm (h) x 9 cm (d))) at two locations within the arena: row 4, column 2 and row 4, column 6 (Figure 2C).
- Keep the objects used for the intra-arena cues and their position constant throughout the experiment but clean them daily with 70% ethanol.
- Set up the 3D extra-arena cues: position distinctive landmarks (e.g., patterned spherical lantern (40 cm (d)); red star lantern (60 cm (w)); blue lantern (70 cm (h) x 35 cm (w))) and patterns around the perimeter of the event arena-which is positioned in the center of the experimental room (Figure 2D).
- Keep the objects used for the extra-arena cues and their position constant throughout the experiment.
- Experimental and control rooms are either one room separated into two parts or two adjacent rooms separated by a curtain or door and are required for this experiment.
- Black boxes
- To enable the animals' access to the arena, build four identical black boxes from black Plexiglass (length: 30 cm, width: 25 cm, height: 35 cm per box; Figure 2E). Each black box should have a remote-controlled sliding door on one-length surface. This will give the experimenter control over the rats' entry to the arena.
NOTE: The black Plexiglass creates a dark interior, which is preferred by the rats over the bright environment of the open-field event arena. - Place these black boxes midway along each of the four walls of the arena. These black boxes will be identified by their location relative to the top of the live video feed-captured by the camera and received by the computers in the control room-using the cardinal points North (top), East (right), South (bottom), and West (left) of the event arena.
- Allow rats to enter the arena from one of the three black boxes, which is referred to as startbox (e.g., East, South, and West; Figure 2A, orange rectangles). Use the remaining black box (e.g., North; Figure 2A, blue rectangle) as a home-base, which the rats will enter to eat the food reward (i.e., pellets) they retrieve from the arena.
NOTE: Any black box location (i.e., North, East, South, West) can be designated as the home base, but it must be kept constant throughout the entire experiment: the stability of its location is critical for the successful encouragement of allocentric spatial representations. - In the startboxes and home-base, put two small, transparent, flat-based wells, one for water and one for food pellets (in the case of the home-base, this is used for reward in the habituation stage only) and place sawdust in each startbox and the home-base.
- To enable the animals' access to the arena, build four identical black boxes from black Plexiglass (length: 30 cm, width: 25 cm, height: 35 cm per box; Figure 2E). Each black box should have a remote-controlled sliding door on one-length surface. This will give the experimenter control over the rats' entry to the arena.
- Sandwells
- Use transparent acrylic plastic, with an inner diameter (d) of 6 cm and a total depth (h) of 6 cm to make sandwells used to conceal the food reward that the rats retrieve (i.e., locate, dig up and take to the home-base to eat). Insert a spherical, perforated plastic bowl 4 cm from the top. Insert the sandwells into the adapted tiles within the arena.
NOTE: The plastic bowl creates an accessible part (6 cm (d) x 4 cm (h)) for rewarded pellets, which the rats have access to, and an inaccessible section (6 cm (d) x 2 cm (h)), to which the rats do not have access (Figure 3A,B). - In the rewarded sandwells, for both encoding and recall choice trials, place four 0.5 g pellets in the accessible section and eight food pellets in the inaccessible section (Figure 3C). In the non-rewarded sandwells, place twelve pellets in the inaccessible section (Figure 3C).
NOTE: Both the rewarded and non-rewarded sandwells contain a total of 12 pellets and are filled with specially prepared sand, which conceals the pellets in the sandwells. - Fill sandwells with a mixture of sand and masala powder (2.5 g masala/2.5 kg sand) to mask any odor emanating from the food pellets. Freshly prepare the sand/masala mixture at the start of every session (Figure 3D).
NOTE: Steps 2.3.2 and 2.3.3 are designed to mask any olfactory artifacts emanating from the sandwells during encoding and choice trials. This will ensure that the rats' search for the correct sandwell location, and their resulting task performance, is guided solely by their memory of the location where food was dug up, and not by any odor cue emanating from the rewarded sandwell, which could reveal the presence of food-reward. - During a probe trial, which tests the rats' memory for the location of the single sandwell previously rewarded (i.e., contain food pellets), make all five sandwells present within the arena as non-rewarded (i.e., no food pellets are available in the accessible section); including the correct sandwell location.
NOTE: All sandwells present within the arena contain the same number of pellets (n = 12) in their inaccessible section.
- Use transparent acrylic plastic, with an inner diameter (d) of 6 cm and a total depth (h) of 6 cm to make sandwells used to conceal the food reward that the rats retrieve (i.e., locate, dig up and take to the home-base to eat). Insert a spherical, perforated plastic bowl 4 cm from the top. Insert the sandwells into the adapted tiles within the arena.
- Overall set-up and software
- Maintain illumination of the experimental room at a moderate level of brightness using wall-mounted halogen lamps (115-125 lux), and maintain a room temperature between 19 to 23 °C.
- Install a charge-coupled device camera above the event arena in the experimental room to record and monitor the rat movements and behavior (Figure 4A). The camera provides a live feed to the adjacent control room for both custom video capture and the custom computer software (developed by P. A. Spooner, University of Edinburgh).
- Monitor the rats' movements using the custom computer software used to time the rats (Figure 4B). This program controls the door of each black box, allowing the experimenters to remotely manage the rats' access to and from the arena from their adjacent control room. Record each animal's latency to locate the correct sandwell and time spent digging at each sandwell during a choice and probe trial.
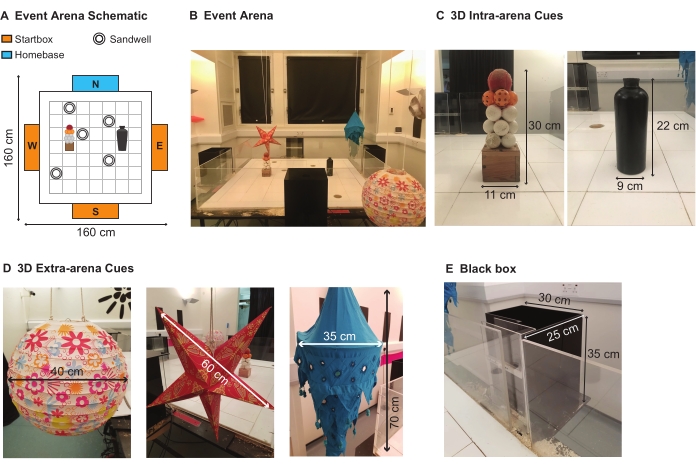
Figure 2: The event arena and cues. (A) Schematic showing the event arena (Abbreviations: N= North, E= East, S= South, W= West). (B) The event arena with intra- and extra-arena cues. (C) The two 3D intra-arena cues (left to right): golf ball stack and cylindrical black bottle. (D) Several 3D extra-arena cues (from left to right): patterned spherical lantern; red star lantern; blue lantern. (E) One of four black boxes is positioned midway along each event arena wall. Three of these black boxes serve as start-boxes, which provide a starting position for the rats at the start of each trial. The fourth black box is a home-base where rats consume the food reward that they retrieve from the arena. Please click here to view a larger version of this figure.
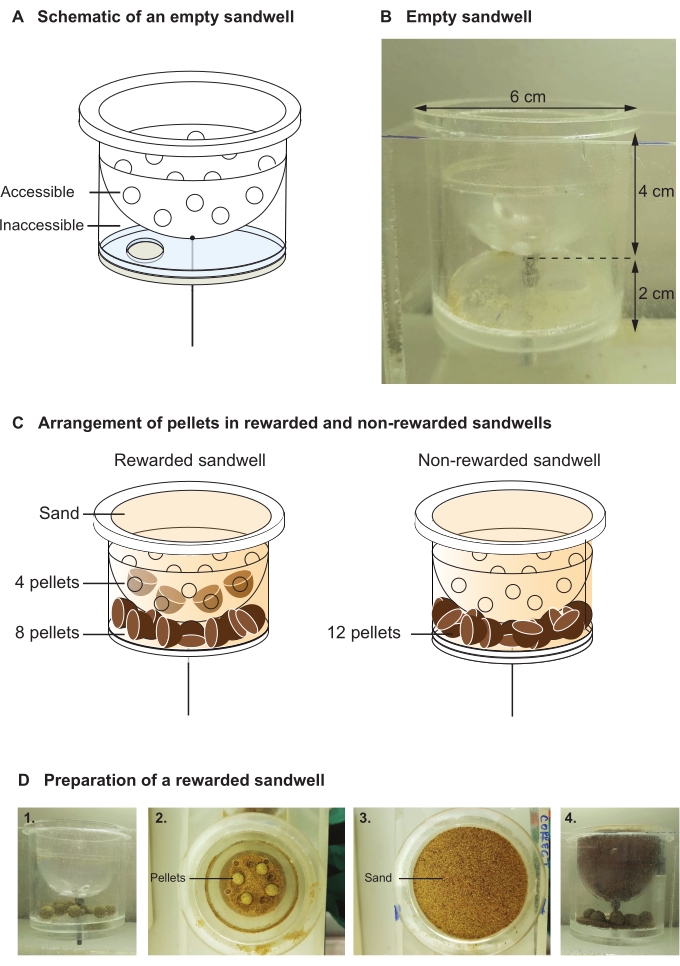
Figure 3: Sandwells. (A) Schematic showing an empty sandwell with the accessible and inaccessible sections labeled. (B) An empty sandwell with an accessible section and inaccessible section. (C) Schematic illustrating the pellet arrangement in a rewarded (left) and non-rewarded (right) sandwell. Both the rewarded and non-rewarded sandwells contain a total of 12 pellets and are filled with specially prepared sand, which conceals the pellets in the sandwells'. (D) Series of photographs showing the preparation of a rewarded sandwell, including the correct placement of the pellets in the accessible section (step 1-4). Please click here to view a larger version of this figure.
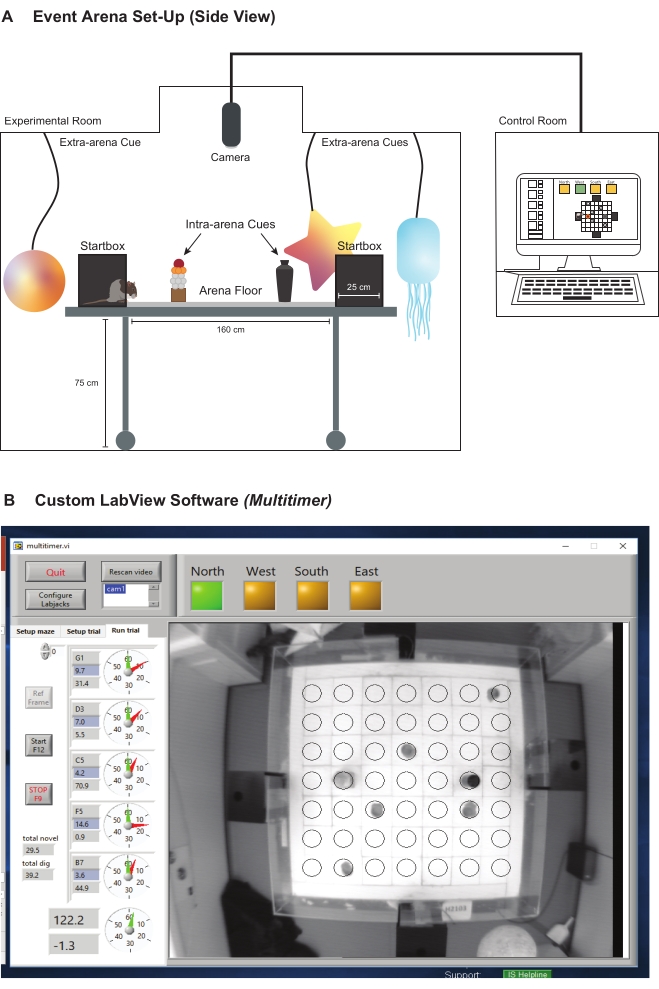
Figure 4: The experimental setup of the event arena. (A) Schematic showing the experimental setup of the experimental and control rooms. (B) Screenshot showing a live feed of the experimental room viewed through the custom computer software. The custom computer software allows the experimenters to control the startbox doors remotely and provides other measurements. Please click here to view a larger version of this figure.
3. Habituation protocol
NOTE: During habituation, the rats are trained to search for sandwells, dig for a food reward, and explore the event arena.
- Learning to dig for a food reward
- Place a small container filled with sand in an individual cage. For the first session (i.e., H1), add three 0.5 g food pellets just under the sand's surface and place one food pellet on top. Thereafter, place all four pellets under the surface (Figure 5).
- Place each rat in the individual cage with one sandwell, refreshing the sandwell for each rat. Leave the rat in the individual cage until it digs and successfully retrieves all four pellets from the sandwell.
- Habituation Session 1
- Place a rat in one of the startboxes (e.g., South) for 30 s and put a pellet (0.5 g) in the home-base's (e.g., North) small, flat-based well designated for food. The placed food pellet acts as a reward and encourages rats to go to the home-base to eat.
- Open the startbox door (e.g., South). Close the door when the rat leaves the startbox, enters the arena, and begins exploring this new environment.
- If the rat does not leave the startbox within 5 min, intervene in one of the two ways. Either encourage the rat to enter the arena by holding a paintbrush just outside of the startbox door. Once the rat is interested, move the brush further into the arena letting the rat follow it. Once the rat is in the arena and a safe distance from the startbox door, close the startbox door.
- Alternatively, take the rat out of the startbox, close the startbox door, and place the rat in the arena, directly outside the startbox door from which it would have entered. If a rat is unmotivated and does not perform the task well (e.g., leave the startbox unassisted, dig effectively, etc.), check the rat's weight and calculate its free-feeding weight (%).
NOTE: If the free-feeding weight is well above 85%, the rat may not be hungry; in this case, its total daily food allowance (g) may require further restriction. - After the rat has explored the arena for 10 min, open the North black-box door (i.e., home-base). If the rat does not enter within 5 min of the door being opened, take the rat out of the arena, close the door of the North black-box, and place the rat in the home-base.
- After the rat has finished eating the pellet placed in the home-base at the start of the session, return the rat to its home cage.
- Habituation Session 2
- Place a sandwell, with all four food pellets (0.5 g/pellet) buried under the surface of the sand, in the event arena. Change the location of this rewarded sandwell each session hereafter.
NOTE: Having large food pellets (0.5 g), the rats will prefer to carry them to an environment they consider safe (i.e., dark environment) to eat them22. - Place one cue pellet in the flat-based well, designated for food, in the chosen startbox (i.e., East), followed by a rat.
- When the rat finishes eating the cue pellet-after approximately 45 s-open the startbox door (e.g., East).
- Close the startbox door once the rat enters the arena and is a safe distance from the door. If the rat does not come out of the startbox, refer to steps 3.2.3-3.2.4.
- Let the rat search for the first pellet in the sandwell. To successfully retrieve the food reward, it must dig in the single sandwell now present within the arena.
- Once the rat retrieves the first pellet, open the home-base door (i.e., North). The rat should then locate and enter the home-base to eat its reward. If the rat starts to eat the pellet within the arena, gently guide it back to the home-base to eat the pellet.
NOTE: This is critical, as each rat must be encouraged to eat in the home-base; without proper training, they have a tendency to return to the trial's startbox, from which they entered the arena, to eat. - After the rat finishes the first pellet at home-base, allow it to leave the home-base and re-enter the arena to retrieve the second pellet.
- Upon retrieving the second pellet, let the rat locate the home-base again to eat the food reward. Once the rat enters the home-base, close the door of the North black box.
- After the rat finishes eating the second pellet in the home-base, gently remove it from the home-base and return the rat to its home cage.
- Place a sandwell, with all four food pellets (0.5 g/pellet) buried under the surface of the sand, in the event arena. Change the location of this rewarded sandwell each session hereafter.
- Sessions 3-7
- Repeat habituation session 2 (steps 3.3.1-3.3.9) five times, burying the pellets in the sandwell deeper with each session (Figure 5). By the end of habituation, encourage all the rats to run quickly to the rewarded sandwell present within the arena, successively collecting the available food pellets and carrying them back to the home-base to eat.
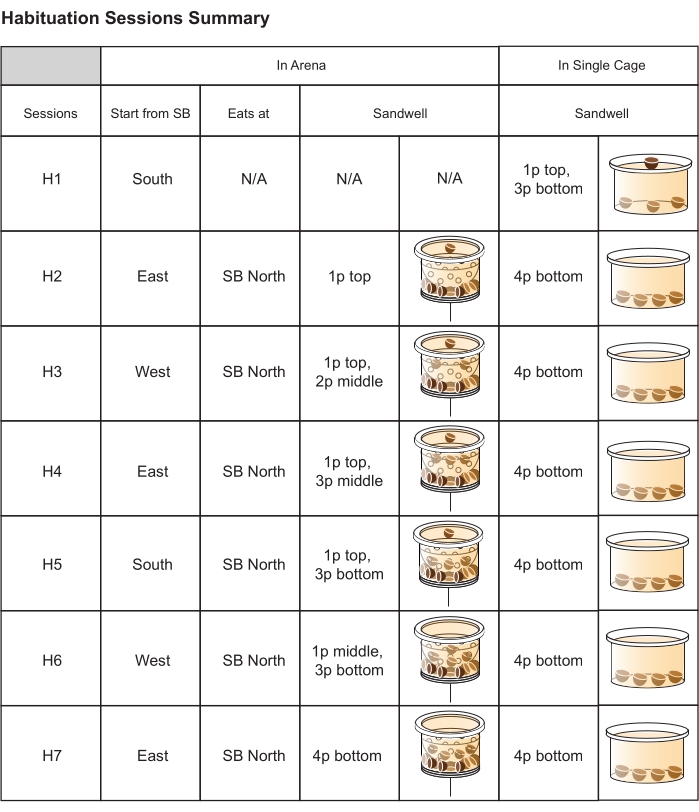
Figure 5: The design of habituation sessions. From left column to right column: the habituation session (H1-H7); the startbox used for each session (e.g., H1: South startbox (SB)); the location where rats are required to eat their food reward (i.e., North home-base); the position of the accessible pellets in the rewarded sandwell (in both written and illustrated form; p = pellet), which will be placed in each session's designated sandwell location; the position of the pellets in the flat-based sandwell in the single cage (in both written and illustrated form), which aims to promote digging behavior and strengthen the rats' association between digging in a sandwell and receiving a food reward. The last two columns refer to sandwells in the single cages (outside the arena). Abbreviation: N/A= not applicable Please click here to view a larger version of this figure.
4. Main training protocol
NOTE: Each main training session consists of two memory encoding trials (E1, E2) followed, after a short time delay (~30 min), by one recall choice trial (C1). During all trials, rats are required to successively retrieve two pellets from the rewarded sandwell. After locating each pellet, the rats should locate and enter the home-base to eat this food reward. The location of the correct (i.e., rewarded) sandwell is counterbalanced across sessions for all rats (Figure 5).
- Counterbalancing measures
- Carefully counterbalance the sequence of the sandwell locations and startbox order used across sessions (Figure 6). Before each session, prepare the location map (Figure 6A); decide the correct sandwell location for each rat, which must notably change across sessions (Figure 6B); and create the counterbalance sheet (Figure 6B,C) and recording sheet (supplementary figure 1).
- Produce three sandwell sets, with five sandwells per set (Figure 6C). Each set requires five sandwells because five sandwell locations (one correct and four incorrect) are used in the arena during each recall choice trial, and three sandwell sets are required so that the sandwells used for each trial can be alternated within every session.
- During each session, use one sandwell set for a rat's encoding trials (Figure 6B,C; Encoding 1: Set 1, Well A; Encoding 2: Set 1, Well B) and another, different sandwell set (Figure 6B,C; Recall Choice: Set 2, Well C) for their recall choice trial.
- Within each session, use a different combination of sandwell sets for each rat (Figure 6B,C), and across sessions, alternate the sandwell set combinations used for each rat.
- Encoding trials
- Put a rewarded sandwell in the correct location according to the location map and the counterbalance sheet (Figure 7). Never use the intra-arena cue locations, the center tile, or the three tiles directly in front of the four startboxes as a sandwell location.
- Place one pellet, followed by a rat, in the startbox (e.g., East) designated for encoding trial 1 (E1); the pellet will act as a cue for the task. Allow enough time (~30 s) for the rat to eat this cue pellet before starting the trial.
- Press the on-screen Start button to record the trial on the in-house video capture system.
NOTE: It is important to maintain a record of the rats' encoding trials for (1) research transparency (i.e., raw evidence of each animal's task performance), (2) re-scoring, and (3) future reference (i.e., to explore and collect the data for other performance measures). - Open the door of the startbox remotely using the custom computer software (Figure 4B).
- Start the timer in the custom computer software when the rat enters the arena and close the startbox door.
- Give the rat 200 s to look for the correct sandwell, dig, and retrieve its first pellet. If the rat has still not located the correct sandwell or its first pellet after 200 s, retrieve one of the pellets from below the sand and place it on top. If the rat fails to visit the correct sandwell and retrieve this pellet after another 200 s, use a brush to guide it gently to the correct sandwell.
- Once the food reward has been found, the rat should carry it to the home-base (e.g., North black box) and eat it once inside. If the rat does not locate and enter the home-base, and chooses instead to eat its first pellet within the arena, quickly take the rat from the arena and place it in the home-base.
- After eating its first pellet in the home-base, let the rat enter the arena from the home-base and locate its second pellet from the correct sandwell.
- After retrieving its second pellet, let the rat locate and enter the home-base to eat it.
- Close the home-base door once the rat is safely inside and give it sufficient time to eat the second pellet.
- Stop the custom video capture recording and timer on the custom computer software. Press the on-screen Stop button on the custom video capture software. Then, click on the Stop button on the on-screen timer on the custom computer software.
- While the rat is eating, wipe the arena floor with a cloth soaked in 70% ethanol solution. Do this between every trial.
- Prepare the correct sandwell for encoding trial 2 (E2) and place it in the correct location within the event arena.
- Take the rat from the home-base and put it in the startbox (e.g., West) designated for E2.
NOTE: The use of an alternate startbox is critical for the effective encouragement of the rats to use only an allocentric spatial solution to perform the task, as the rats cannot rely on a static viewpoint of the arena or follow their previous path to successfully locate the correct sandwell. Instead, they need to attend to the intra- and extra-arena cues, which promotes allocentric encoding. - Repeat steps 4.3.2 to 4.3.12, and then return the rat to its home cage.
- Recall choice trial
NOTE: Each rat's recall choice trial is run 30-40 min after the second encoding trial (E2) and presents the rats with an arena containing five sandwells.- Place the rewarded sandwell in the correct location assigned for the session, while the four unrewarded sandwells are placed in the four incorrect locations allocated for the session and rat in question (Figure 8A). The sandwell location map for the five sandwells in the recall choice trial is altered for each session and counterbalanced across sessions.
- Put the sandwell containing four accessible pellets in the correct location. Put four additional sandwells-each unrewarded and containing no pellets in the accessible section-in the incorrect locations set by the session's sandwell location map.
- Put the cue pellet and rat in the startbox for the recall choice trial (e.g., South). Ensure that this starting location (e.g., C1: West) is different from those used in the two encoding trials (e.g., E1: East, E2: South).
- Start recording the trial using the in-house video capturing system.
NOTE: It is important to maintain a record of the rats' recall choice trials for (1) research transparency (i.e., raw evidence of each animal's task performance, which could be submitted as part of a paper's supplementary material), (2) re-scoring, and (3) future reference (i.e., to explore and collect the data for other performance measures). - On the custom computer software, select the timers matching the sandwells (sandwell timers) to be used on this particular session (Figure 4B).
- Once the rat has eaten the cue pellet (~30 s), open the door using the custom computer software. When the rat leaves the startbox, close the startbox door and start the timer on the custom computer software.
- When the rat digs in a sandwell, click the on-screen Sandwell icons to record the time spent digging in each sandwell. Continue to record the rat's dig time in each sandwell visited until the end of the recall choice trial.
- Allow the rat to then locate and enter the home-base to eat this food reward.
- Use the same procedure as that given for the encoding trials (see step 4.2) for the retrieval of the second pellet from the correct sandwell in the recall choice trial.
- Do not click and record the rat's dig time in each sandwell visited during its search for the second pellet. Only record the order of the sandwells (using the number 1-5 allocated to each sandwell location) visited before the rat successfully locates the rewarded sandwell and retrieves its second pellet. This requires concentration.
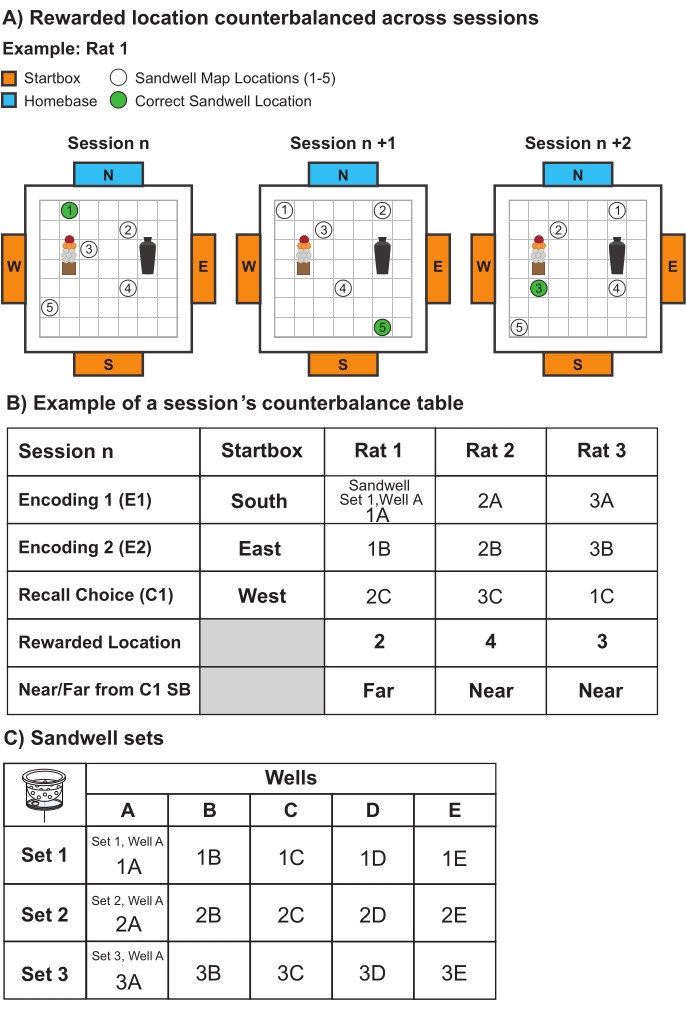
Figure 6: Representative counterbalancing. (A) Schematic illustrating how the sandwell location map and correct sandwell location encountered by the rats (e.g., Rat 1) changes across the sessions. (B) Example of a counterbalance table for one session (e.g., Session 1). A different startbox is used for each trial within a single session (i.e., encoding trial 1 (E1) started from the South startbox (SB)), but their order of use was the same for each animal (e.g., Rat 1-3). The sandwells used for the correct location (e.g., location 2, 4, 3) and their associated sets, used in full during the recall choice trial, were counterbalanced across each session's trials (e.g., encoding 1, encoding 2, recall choice) and the animals performing the task (e.g., Rat 1-3). (C) Table outlining the sandwell sets counterbalanced within and across sessions. There are 15 sandwells in total and three sets (set 1-3) of sandwells, each containing five wells (A-E). Each rat uses different wells in each encoding and recall choice trial. For example, as mentioned in Figure 6B, Rat 1 will use Sandwell 1A in encoding trial 1, Sandwell 1B in encoding trial 2, and Sandwell 2C in the recall choice trial. Please click here to view a larger version of this figure.
5. Recall probe test
- Use the same set-up as that for a recall choice trial, except that there are no accessible pellets in any of the five sandwells, including the session's previously rewarded, correct sandwell location (Figure 8A,B).
NOTE: As in the recall choice trial, all five sandwells are available during the probe test and the rats are free to dig in any sandwell(s) they choose; however, none of the sandwells contain an accessible food reward-instead, all 12 pellets are present in the inaccessible section of each sandwell (Figure 3C). - Put the five sandwells containing no accessible pellets in the arena at the locations provided in the session's sandwell map (Figure 8).
- Put the rat in the startbox with a cue pellet. Use the starting position not employed in either of the same session's two encoding trials.
- Set the sandwell timers on the custom computer software to correspond with the session's sandwell map (Figure 4B). Ensure the sandwell timers set on the custom computer software correctly correspond to the session's sandwell map.
- Start recording the probe trial in the in-house video capturing system.
NOTE: It is important to maintain a record of the rats' recall probe trials for (1) research transparency (i.e., raw evidence of each animal's task performance, which could be submitted as part of a paper's supplementary material), (2) re-scoring, and (3) future reference (i.e., to explore and collect the data for other performance measures). - Once the rat has finished the pellet, open the startbox door remotely using the custom computer software (Figure 4B).
- Once the rat has entered the arena and is a safe distance from the door, close the startbox door and start the timer in the custom computer software.
- Record the rat's dig time and latency to each of the sandwells visited during the 120 s probe trial, by clicking on each sandwell visited and holding for as long as the rat continues to dig. This 120 s countdown commences when the rat digs in the first sandwell.
- Record the dig times and latencies at 60 s and 120 s by taking a screenshot of the custom computer software at the 60 s and 120 s time mark.
- After the 120 s probe trial has elapsed, put three pellets in the correct sandwell (i.e., the location of the rewarded sandwell in the encoding trial) to prevent memory decline. The rat is required to retrieve two of these three pellets. Once a pellet is retrieved, the rat is required to locate and enter the home-base to eat it.
- Press the on-screen Stop button on the custom computer software after the 120 s probe test. Clear the file name and note only the rat's latency to retrieve the first and second pellet placed in the now-rewarded, correct sandwell location.
NOTE: Preferential digging at the correct location is used as an index of memory: good memory for the everyday event, experienced in the encoding trials (i.e., encountering the session's correct sandwell location), is indicated by a greater time spent digging in the correct location than the average time spent digging in the incorrect locations. - Schedule a recall probe test at the beginning of the training to check whether the performance is at chance level. Thereafter, schedule probe tests at specific intervals (e.g., every sixth session), or schedule only when the rats reach a stable task performance: to warrant a probe test, their average performance index (%) needs to be 60% or above for three consecutive sessions. The average performance index is defined in step 7.3.1.
6. Non-encoding control test
NOTE: A non-encoding trial is a control measure used to determine whether the rats are using olfactory artifacts, rather than their memory of the correct sandwell location, to perform the task. As the name suggests, the 'non-encoding control test' means that there are no encoding trials performed prior to the recall choice trial; only the recall choice trial is conducted. The expectation is that without being permitted to encode the location of the everyday memory event, the rats' performance in the choice trial will be at chance level. If this is not the case, and the rats perform well in the non-encoding control test, a re-design of the sandwells, and their accessible and inaccessible compartments may be required.
- Perform a recall choice trial as described in section 4.3 (steps 4.3.1 to 4.3.10).
7. Performance measurement
NOTE: Several parameters are measured and Supplementary Figure 1 shows an example datasheet.
- Choice of sandwells
NOTE: Choice is defined as the number of sandwells that rats dig in, up to and including the correct sandwell, during the recall choice and recall probe trials. The maximum possible value of the choice is 5, as there are five sandwells in total.- During each trial of the experiment (recall choice trial and recall probe test), determine the number of choices made by a rat: whether it places its front paw(s) on or into a sandwell. If a rat ran past, or merely sniffed quickly in the vicinity of a sandwell, this is not considered a choice.
- In rare cases, when it is difficult to tell from the video monitors whether the rats make a choice (as defined above), check at the end of the trial, whether there are any traces of digging: that is if the sand is displaced around the sandwell(s). If there is evidence of digging, however slight, consider this a choice. Pausing at a sandwell and not digging is considered only a visit and should not be counted as a choice.
- Errors
NOTE: Error is defined as the number of incorrect sandwells (unrewarded) that rats visit before locating the correct sandwell. Choice is defined as the number of sandwells that rats dig in, up to and including the correct sandwell, during the recall choice and recall probe trials. The maximum number of errors is four as there are five sandwells in total.- Calculate error using the following formula:
Error = (Choice – 1) - When a rat re-visits the incorrect sandwell, do not count this as another error as the maximum number of errors is four due to there being five sandwells in total.
- Calculate error using the following formula:
- Performance index (PI)
NOTE: Performance index is defined as the number of errors made before the rats locate the correct sandwell in a recall choice trial. With five sandwells, up to four errors can occur. The chance level among five sandwells is therefore two errors (i.e., 50%).- Calculate performance index using the following formula:
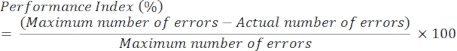
- When a rat re-visits the incorrect sandwell, do not count this as another error as the maximum number of errors is four due to there being five sandwells in total.
- Calculate performance index using the following formula:
- Latency
NOTE: Latency is defined as the time that elapses before digging commences at the correct sandwell(s).- Measure latency from when the rat leaves the startbox until it reaches the correct sandwell. Monitor and record latency using the custom computer software.
- Dig time
- Measure the rats' dig time in each sandwell (both correct and incorrect sandwells) in the recall probe trial.
NOTE: Good memory for the everyday event is defined by the rats' digging in the correct sandwell (n = 1) for a greater proportion of the 120 s probe trial than the average time they spend digging in the incorrect sandwells (n = 4). - Calculate the correct and incorrect using the following formulae:
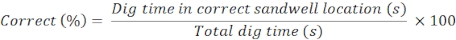
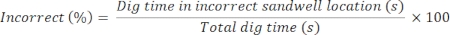
- Measure the rats' dig time in each sandwell (both correct and incorrect sandwells) in the recall probe trial.
8. Avoidance of unintended bias
NOTE: The following control measures are implemented throughout the protocol to ensure the reproducibility and reliability of this everyday memory task.
- Counterbalance the sandwell locations across sessions. This avoids garnering any reward bias to a specific side of the event arena.
- Counterbalance the sandwell sets, as well as the sandwells within these sets used in the correct position, across sessions and rats within each session. This discourages the rats from attempting to follow any residual odor trial lingering from the trials of the preceding rat(s).
- Wipe the floor of the event arena between every trial with a cloth soaked in 70% ethanol solution; this will prevent the path of a previous rat(s) from influencing subsequent task performance.
This stable home-base protocol has been used to successfully train rats to learn this everyday memory task using allocentric representations. There are two important elements in this protocol. First, animals start from different black boxes (e.g., East, South, and West) within and between sessions (Figure 7A). There are two encoding trials and one recall choice trial per session (or probe trial instead of the choice trial in some cases), all starting from an alternate startbox. This encourages the animals to attend to both the intra- and extra-arena cues upon entering the arena. It is not possible to solely rely on idiothetic path integration to perform the task, as different paths are required from each startbox to reach the correct sandwell location. Second, the animals readily learn to eat the food reward pellets in the North-located, stable home-base. During both the encoding trials and recall choice trials, the rats dig and collect their food reward, and then, very naturally, run back to the North home-base to eat the first pellet (Figure 7A). After they finish the first pellet, they come out from the North box and look for the second pellet. This is a different path to that of the first pellet and the rats have to re-orient themselves to successfully relocate the correct sandwell. Again, this encourages the animals to consider the intra- and extra-arena cues and promotes the use of allocentric representations.
Memory formation: We first examined whether rats could achieve a stable performance using this home-base protocol. We found that the rats acquired a good, above-chance performance in this everyday memory task within 16 sessions (Figure 7B). The performance index for recall choice trials peaked at around 80%: a level of performance comparable to previous everyday memory protocols, which did not use a stable home-base14,15,17. While 80% may not seem as good as the 90% and above reached in two-alternative forced-choice tasks11, bear in mind that this is a five-alternative choice task. The stability of performance across sessions was also impressive (typically <7%). We also took steps to ensure that this level of performance was based on the rats' memory of the everyday event encoded during the two encoding trials through the use of two non-encoding control tests. If the animals were artifactually relying on cryptic olfactory cues, their performance would be above chance level (50%); however, if the animals were instead relying on their memory of the everyday event, encountered during the two encoding trials, their performance would be poor and fall to chance level (50%). When we conducted these non-encoding control tests, the rats' task performance fell to chance level (Figure 7B). From this result, we conclude that the rats were relying on their memory of the everyday event to successfully perform the task.
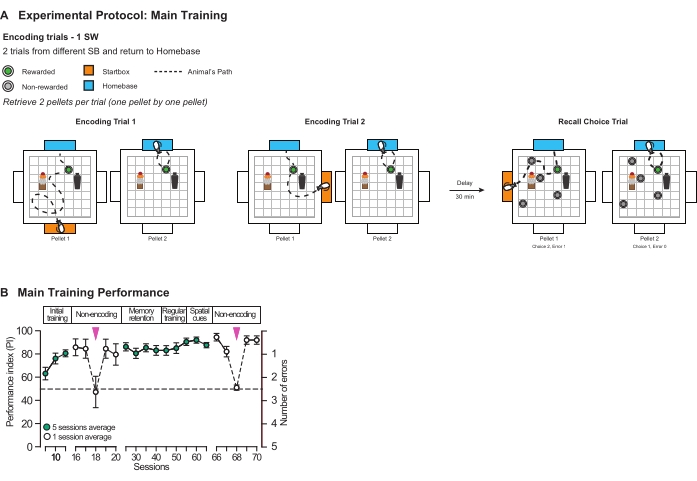
Figure 7: Main training protocol and performance index. (A) Schematic outlining the experimental protocol for the everyday memory task's main training. During each main training session, two encoding trials (encoding trial 1 and encoding trial 2) were first performed. During each encoding trial, the rats were trained to retrieve two pellets successively (one pellet by one pellet) from the single, correct (i.e., rewarded, green) sandwell located within the event arena. Each encoding trial began from a different startbox (orange). To retrieve the food reward, the rats left the startbox (e.g., South) and located the correct sandwell (green). Once the rats retrieved the food reward from the correct sandwell, they located and entered the home-base (North, blue) to eat the food pellet. After the rats retrieved the second pellet in encoding trial 2, they experienced a short 30 min delay, followed by a recall choice trial. Starting at a different startbox during encoding trials, the rats encountered an arena where multiple incorrect sandwells (gray) and one correct sandwell (green) were now present. (B) Graph showing the rats' (n = 17) acquisition data for this stable home-base task. The rats achieved a consistently good task performance by session 16 (ANOVA), which was maintained until session 70 (above chance, t-test, p < 0.05 or better). Two non-encoding control trials were performed at the start (session 18) and end (session 68) of the main training program (pink arrows). In the absence of the encoding trials, the rats performed poorly: their average performance index (%) fell to chance level (50%, t-test, p > 0.05). Data are mean ± SEM. This figure has been modified from Broadbent et al18. Please click here to view a larger version of this figure.
Limited retention over time: After establishing that the rats could successfully remember spatially located events in this allocentric protocol, we tested whether they displayed overnight forgetting, which is characteristic of episodic-like everyday memory (Figure 8A). We tested the rats with two retention delays of 24 min and 24 h – and used a within-subject paradigm, in which every animal experiences every condition. The rats spent significantly more time digging at the correct location after the 24 min delay than the 24 h delay (t(7) = 2.85, p < 0.05) (Figure 8B). This delay-dependent forgetting – an essential feature of everyday memory – ratifies the effectiveness of our task: everyday memory can be effectively modeled by this protocol, as memory decays over the course of 24 h.
Allocentric encoding: Next, we examined whether the rats rely on the intra- and extra-arena cues to successfully perform this memory recall task. After the two encoding trials and before the recall probe trial, a curtain was placed around the arena to remove sight of all intra- and extra-arena cues, followed by the anticlockwise rotation of the arena by 45°. In the probe trial, where all cues (intra- and extra-arena) were hidden or removed, the rats' task performance significantly declined to chance level (t(7) = 3.37, p < 0.05) (Figure 8C,D). This result strongly suggests that use of a home-base effectively encourages the rats to employ an allocentric spatial strategy. Furthermore, in the original study, we also performed inter-experimenter correlations to confirm that all the experimenters involved with running this behavioral task recorded the rats' performance similarly18.
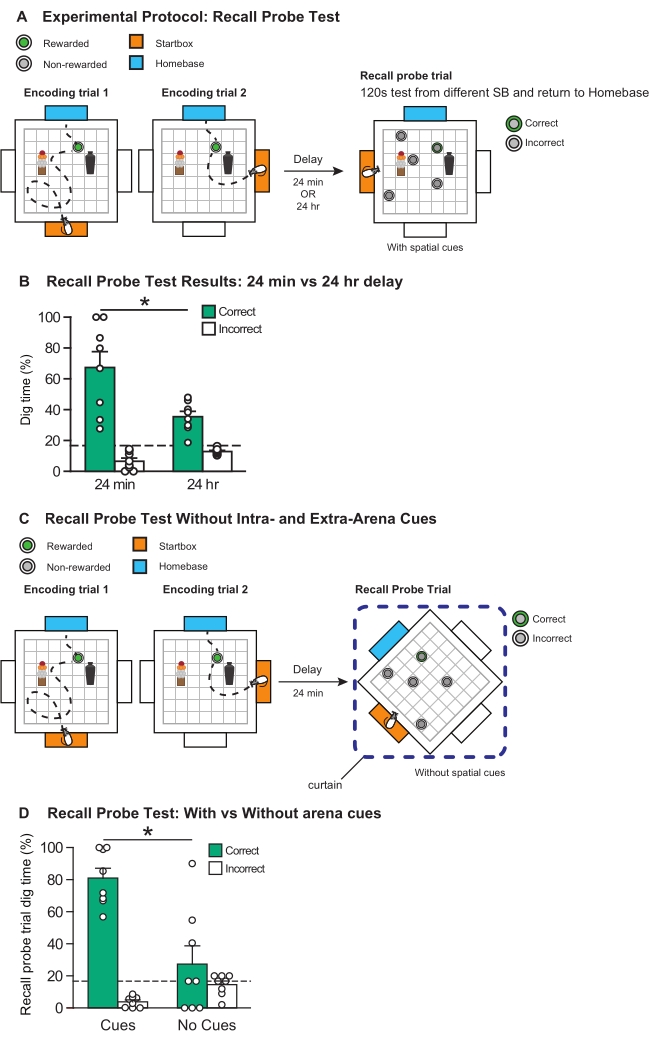
Figure 8: Recall probe test protocols and results. (A) Schematic illustrating the experimental protocol for a recall probe test session. Rats were trained in two encoding trials, and after a delay of either 24 min or 24 h, were presented with five sandwells. (B) Graph showing the characteristic delay-dependent decay of everyday memory. After a delay of 24 min, the rats' memory for the encoded event was significantly above chance level (t (7) = 2.92, p < 0.05) and significantly different from 24 h (t (7) = 2.85, p < 0.05); *p < 0.05. Data are mean ± SEM. Individual data points are also shown. (C) Schematic showing the protocol for the probe test used to assess spatial strategy, where all intra- and extra-arena cues were hidden behind curtains or removed, and the arena was rotated by 45°. The encoding trials were run identically to those performed in a normal probe test (Figure 8A, i.e., all environmental cues are present). The delay between encoding trial 2 and the recall probe trial was 24 min, and all environment cues were removed for the recall probe trial. (D) Graph showing the results of probe test with and without the event arena's environmental cues. When the intra- and extra-arena cues were removed, the rats performed poorly, digging in the correct sandwell for a significantly lower proportion of the 120 s probe trial than observed when all environment cues were present (t (7) = 3.70, p < 0.05). *p < 0.05. Data are mean ± SEM. Individual data points are also shown. This figure has been modified from Broadbent et al18. Please click here to view a larger version of this figure.
Supplementary Figure 1: Example of a recording sheet used to record a single rat's performance during encoding trial 1, encoding trial 2, and the recall choice/probe trial of a single session. Please click here to download this File.
