

Summary
نحن نقدم منهجية لتحديد متطلبات التلقيح من المشمش(Prunus أرمينياكا L.) أصناف الجمع بين تحديد الذاتي (في) التوافق بواسطة المجهر fluorescence مع تحديد النمط الجيني S من قبل تحليل PCR.
Abstract
يتم تحديد عدم التوافق الذاتي في الوردية من قبل نظام عدم التوافق الذاتي (GSI) الذي يتم التحكم فيه بشكل رئيسي من قبل loallelic S. في المشمش ، وتحديد الذات وفيما بين (في) علاقات التوافق هو على نحو متزايد أهمية ، منذ الإفراج عن عدد كبير من أصناف جديدة قد أدى إلى زيادة أصناف مع متطلبات التلقيح غير معروف. هنا، نحن وصف منهجية التي تجمع بين تحديد الذاتي (في) التوافق عن طريق التلقيح باليد والمجهر مع تحديد من S-نوع جيني من قبل تحليل PCR. لتحديد التوافق الذاتي (في) ، تم جمع الزهور في مرحلة البالون من كل أصناف في الحقل ، ملقحات يدوي في المختبر ، ثابتة ، وملطخة باللون الأزرق الأنيلين لمراقبة سلوك أنبوب حبوب اللقاح تحت المجهر الفلوري. 10- ولإقامة علاقات عدم التوافق بين الأصناف، استُخرج الحمض النووي من كل صنف من أوراق الشجر الصغيرة، وتم تحديد الـS-alleles بواسطة PCR. ويسمح هذا النهج بإنشاء مجموعات عدم التوافق وتوضيح علاقات عدم التوافق بين الأصناف، مما يوفر معلومات قيمة لاختيار الملقحات المناسبة في تصميم البساتين الجديدة واختيار الآباء المناسبين في برامج التربية.
Introduction
عدم التوافق الذاتي هو استراتيجية من النباتات المزهرة لمنع التلقيح الذاتي وتعزيز outcrossing1. في Rosaceae، يتم تحديد هذه الآلية من قبل نظام عدم التوافق الذاتي Gametophytic (GSI) Sالتي يتم التحكم فيها أساسا من قبل متعدد الأراضي2. في الاسلوب ، جين RNase ترميز S - stylar المحدد ، RNase3، في حين أن بروتين F - مربع ، الذي يحدد S -حبوب اللقاح المحدد ، هو مقنن من قبل SFB الجينات4. يحدث التفاعل الذاتي عدم التوافق من خلال تثبيط نمو أنبوب حبوب اللقاح على طول النمط منع الإخصاب من البويضة5،6.
في المشمش ، وقد حدث تجديد varietal في جميع أنحاء العالم في العقدين الماضيين7،8. هذا التقديم لعدد هام من أصناف جديدة، من مختلف برامج التربية العامة والخاصة، أدى إلى زيادة أصناف المشمش مع متطلبات التلقيح غير معروف8.
وقد استخدمت منهجيات مختلفة لتحديد متطلبات التلقيح في المشمش. في مجال، الذاتي (في) التوافق يمكن أن تنشأ عن طريق التلقيحات التي تسيطر عليها في الأشجار في قفص أو في الزهور emasculated وتسجيل نسبة الفاكهة في وقت لاحقمجموعة 9،10،11،12. وبالإضافة إلى ذلك، وقد أجريت التلقيحات التي تسيطر عليها في المختبر من قبل شبه في ثقافة الجسم الحي من الزهور وتحليل سلوك أنبوب حبوب اللقاح تحت المجهر الفلوريسنس8،13،14،15،16،17. في الآونة الأخيرة، سمحت التقنيات الجزيئية، مثل تحليل PCR وتسلسله، بتوصيف علاقات عدم التوافق على أساس دراسة جينات RNase و SFB 18،19. في المشمش، ثلاثة وثلاثين S-أليليس تم الإبلاغ عن (S1 إلى S20، S22 إلى S30، S52، S53، Sv، Sx) ، بما في ذلك أليل واحد ذات صلة بالتوافق الذاتي (Sc)12،18،,2121،22،23،24. حتى الآن، 26 مجموعات عدم التوافق قد تم stablished في هذا النوع وفقا ل S-genotype8،,9،,17،,25،,26،,27. أصناف مع نفس S-alleles هي inter-incompatible، في حين أن أصناف مع واحد على الأقل مختلفة S-أليل، وبالتالي، المخصصة في مجموعات مختلفة غير متوافقة، هي inter-compatible.
لتحديد متطلبات التلقيح من أصناف المشمش، ونحن وصف منهجية التي تجمع بين تحديد الذاتي (في) التوافق بواسطة المجهر الفلوريس مع تحديد من S-الجيلية من قبل تحليل PCR في أصناف المشمش. ويسمح هذا النهج بإنشاء مجموعات عدم التوافق وتوضيح علاقات عدم التوافق بين الأصناف.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. الذاتي (في) تحديد التوافق
- تذوق الزهور في الحقل. فمن الضروري لجمع الزهور في مرحلة البالون (الشكل 1A)، المقابلة للمرحلة 58 على مقياس BBCH لامشمش28، لتجنب التلقيح السابق غير المرغوب فيها.
-
التلقيحات الذاتية والمتقاطعة في المختبر
- إزالة anthers من الزهور في مرحلة البالون ووضعها على قطعة من الورق لتجف في درجة حرارة المختبر.
- بعد 24 ساعة، غربال حبوب اللقاح باستخدام شبكة غرامة (0.26 مم) (الشكل 1B).
- Emasculate مجموعة من 30 الزهور في نفس مرحلة البالون لكل التلقيح الذاتي والتلقيح عبر ووضع pistils على رغوة بائع الزهور في الماء في درجة حرارة المختبر (الشكل 1ج).
- تلقيح اليد pistils بمساعدة فرشاة الطلاء مع حبوب اللقاح من الزهور من نفس أصناف 24 ساعة بعد emasculation. وبالإضافة إلى ذلك، تلقيح مجموعة أخرى من pistils من كل أصناف مع حبوب اللقاح من الزهور من ملقحات متوافق كتحكم(الشكل 1دال).
- بعد 72 ساعة، قم بإصلاح مسك هاته الزغر في محلول تثبيتي من حمض الإيثانول/الخليك (3:1) لمدة 24 ساعة على الأقل عند 4 °C29. ثم تجاهل المثبت وإضافة 75٪ الإيثانول ضمان أن العينات مغمورة تماما في الحل. ويمكن حفظ العينات في هذا الحل في 4 درجة مئوية حتى استخدام8،17،30،31،32.
-
تقييم صلاحية حبوب اللقاح من خلال إنبات حبوب اللقاح في المختبر
- لإعداد متوسط الإنبات، الوزن 25 غرام من السكروز، 0.075 غرام من حمض البوريك (H3BO3)و 0.075 غرام من نترات الكالسيوم (Ca(NO3)33.33
- إضافة مكونات المتوسطة في 250 مل من الماء المقطر وتذوب تماما.
- ترسيخ المتوسطة مضيفا 2 غرام من agarose ومزيج من دوامة.
- تحقق من درجة ح H من المتوسطة باستخدام مقياس درجة ح/ و ضبط القيمة إلى 7.0 مع محلول NAة أو HCl.
- اتوملاف الخليط لتعقيم المتوسطة.
- بعد التكلاف التلقائي، يبرد المتوسط ويوزعه على أطباق بيتري في غطاء تدفق معقم.
- تشتت حبوب اللقاح من نفس الأصناف المستخدمة في التلقيحات التي تسيطر عليها في اوسط انبات حبوب اللقاح الصلبة ومراقبة لهم تحت المجهر بعد 24 ح6.
ملاحظة: لتعقيم غطاء تدفق الفاتن، قم بتنظيف السطح بنسبة 70% الإيثانول والتبديل على مصباح الأشعة فوق البنفسجية خلال 10 دقيقة. - يُخزن أطباق بيتري في ثلاجة عند 4 درجات مئوية حتى الاستخدام.
-
ملاحظات الفحص المجهري
- اغسل البيستيل ثلاث مرات لمدة 1 ساعة بالماء المقطر واتركها في كبريتيت الصوديوم بنسبة 5٪ عند 4 درجات مئوية. بعد 24 ساعة، أوتلافها عند 1 كجم/سم2 خلال 10 دقائق في كبريتيت الصوديوم لتنعيم الأنسجة34.
- ضع pistils autoclaved على شريحة زجاجية ، وبمساعدة مشرط ، قم بإزالة trichomes حول المبيض للحصول على تصور أفضل لأنابيب حبوب اللقاح. ثم، الاسكواش pistils مع غطاء الزجاج.
- إعداد 0.1٪ (الخامس / الخامس) aniline وصمة عار الأزرق: مزيج 0.1 مل من الأزرق البيب في 100 مل من 0.1 ن البوتاسيوم الفوسفات ثلاثية الباب (K3PO4). تطبيق قطرة من aniline الأزرق على الاستعدادات لطخة ترسبات الكالوز أثناء نمو أنبوب حبوب اللقاح.
- مراقبة أنابيب حبوب اللقاح على طول النمط عن طريق المجهر مع epifluorescence الأشعة فوق البنفسجية باستخدام 340-380 bandpass و 425 مرشحات طول.
2. استخراج الحمض النووي
- عينة 2-3 يترك في هذا المجال في الربيع. فمن المستحسن أن عينة من الأوراق في مراحل الشباب منذ الحصول على الحمض النووي هو من أعلى جودة ومستويات أقل من المركبات الفينولية مقارنة مع الأوراق القديمة.
- استخراج الحمض النووي الجينومي بعد الخطوات الموضحة في مجموعة متاحة تجاريا (انظر جدول المواد).
- تحليل كمية ونوعية تركيزات الحمض النووي باستخدام مقياس الطيف فوق البنفسجي (260 نانومتر).
3. S-أليلي تحديد الهوية
-
إعداد ردود فعل PCR
- إعداد تخفيف 50 نانوغرام/ميكرولتر في الماء المقطر لكل عينة استخراج الحمض النووي.
- ذوبان من الكواشف PCR ببطء والاحتفاظ بها على الجليد. اترك بوليميراز الحمض النووي في الثلاجة حتى الحاجة.
- إعداد تفاعلات التضخيم باستخدام مجموعات مختلفة من التمهيديات. إنشاء مزيج تفاعل PCR عن طريق الجمع بين المكونات في الجدول 1. دوامة تفاعل PCR مزيج جيد وتوزيع وحدة التخزين المشار إليها لمجموعات مختلفة من التمهيدي لكل بئر من لوحة PCR. ثم، إضافة 1 μL من تخفيف الحمض النووي في كل بئر.
- ضع لوحة PCR في الدراجات الحرارية وتشغيل برنامج PCR المقابلة الموضحة في الجدول 1.
- تحليل شظايا تضخيم. هناك طريقتان مختلفتان أساسا لتحليل شظايا تضخيم البوليميرازيين: الكهرباء الشعرية (CE) مع الإنارة الفلورية أو كما amplicons تصور من الغيلة الغيلة agarose مع التمهيديات غير الموسومة.
-
الشعيرات الدموية الكهربائية
- لإعداد المخزن المؤقت للتحميل، اخلط 35 ميكرولتر من الفورماميد غير المتأين مع 0.45 ميكرولتر من معيار التحجيم المسمى. دوامة الكاشف لخلط جيدا، ومن ثم الاستغناء 35.5 μL في بئر لوحة القارئ.
- أضف 1 ميكرولتر من منتج PCR إلى البئر. وبالإضافة إلى ذلك، إضافة قطرة من الزيوت المعدنية لمنع تبخر المياه.
- إعداد لوحة الفصل مضيفا العازلة الفصل.
- استخدام البرامج التجارية المضمنة مع محلل الجينات (انظر جدول المواد). إنشاء لوحة عينة جديدة وحفظ أسماء العينات لجميع الآبار على لوحة.
- حدد طريقة التحليل. في هذه الحالة، تُسخ العينات عند 90 درجة مئوية لمدة 120 درجة مئوية، وحقن 2.0 كيلو فولت لمدة 30 درجة، ومنفصلة عند 6.0 كيلو فولت لمدة 35 دقيقة.
- أدخل اللوحتين في محلل الجينات. املأ صفيف الشعيرات الدموية بالماء المقطر.
- تحميل براءة اختراع البولي أكرريلاميد الخطية (LPA) هلام. وأخيراً، انقر فوق تشغيل.
-
جل الكهربائية
- إعداد جل agarose 1٪ إضافة 1.5 غرام من الأحياء الجزيئية الصف agarose في 150 مل من 1x TAE (تريس-أسيتات-EDTA) الكهرباء تشغيل العازلة (40 mM تريس، 20 mM حمض الخليك، و 1 mM EDTA في درجة الحموضة 8.0). قم بحل الأاروز عن طريق تسخين الميكروويف لمدة 2-3 دقائق.
- لتصور الحمض النووي، أضف 4 ميكرولتر من بقعة حمض نووي (انظر جدول المواد) واخلط بلطف.
- إضافة مشط هلام، مع آبار كافية للسلالم والضوابط والعينات، في علبة هلام. ثم، صب ببطء مزيج في منتصف علبة هلام وتجنب فقاعات.
- السماح للجل يبرد لمدة 30-45 دقيقة في درجة حرارة الغرفة حتى هلام قد توطدت تماما. إدخال الجل في غرفة الكهرباء، وإزالة مشط هلام وملء الغرفة مع ما يكفي من 1x TAE العازلة لتغطية هلام.
ملاحظة: تحقق من موضع الجل. وينبغي وضع الآبار بالقرب من القطب السلبي لأن الحمض النووي المشحون بشكل سلبي يهاجر نحو الكاثود. - إضافة 5 ميكرولتر من العازلة التحميل (0.1٪ (الخامس / الخامس) الأزرق البروموفينول) لمنتجات PCR ومزيج جيد.
- لتقدير حجم النطاقات، قم بتحميل 5 ميكرولتر من سلم الوزن الجزيئي للحمض النووي (انظر جدول المواد).
- قم بتحميل العينات في الآبار الإضافية للجل.
- مرة واحدة يتم تحميل جميع العينات وسلالم الوزن الجزيئية DNA، تشغيل الجل في 90 الخامس ل1-1.5 ساعة، حتى خط الصبغة الزرقاء هو تقريبا في 75٪ طول الجل.
- تصور العصابات في الترانرلومينات للأحماض النووية.
-
الشعيرات الدموية الكهربائية
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
تتطلب دراسات التلقيح في المشمش استخدام الزهور في مرحلة البالون المتأخر قبل يوم واحد من التحلل(الشكل 1أ). وتعتبر هذه المرحلة الأكثر ملاءمة لكل من جمع حبوب اللقاح وpistil، منذ هياكل الأزهار هي ناضجة تقريبا، ولكن لم يحدث بعد anther dehiscence. وهذا يمنع تدخل حبوب اللقاح غير المرغوب فيها ، ليس فقط من حبوب اللقاح من نفس الزهرة ولكن أيضًا من الزهور الأخرى ، لأن البتلات المغلقة تعوق وصول الحشرات التي تحمل حبوب اللقاح الخارجية. الحبوب حبوب اللقاح هي منخل بسهولة من خلال شبكة غرامة (الشكل 1ب) من anthers dehisced وضعت سابقا على قطعة من الورق لمدة 24 ساعة في درجة حرارة الغرفة أو مع حرارة إضافية طفيفة. وبالمثل، يتم الحصول على pistils من الزهور في مرحلة البالون بعد إزالة بتلات، sepals و stamens بمساعدة ملاقط أو الأظافر (الشكل 1ج). يمكن أن يكون Pistils الذاتي وعبر تلقيح مع فرشاة غرامة (الشكل 1دال).
الزهور hermaphroditic المشمش خمسة sepals الأحمر الداكن، خمس بتلات بيضاء(الشكل 1A)،pistil واحد(الشكل 2أ)و 25-30 stamens. يحتوي الـ pistil على ثلاثة هياكل رئيسية: وصمة العار والأسلوب والمبيض. المبيض لديه اثنين من البويضات، وتخصيب واحد منهم على الأقل مطلوب لإعداد الفاكهة. أثناء التلقيح، الحشرات، النحل أساسا، نقل حبوب اللقاح إلى وصمة العار (الشكل 1أ)، حيث تنبت(الشكل 2ب)في غضون 24 ساعة بعد التلقيح. يتم إنتاج أنبوب حبوب اللقاح من كل حبوب اللقاح germinating، والتي تنمو من خلال هياكل البيستيل للوصول إلى المبيض بعد 3-4 أيام وتخصيب واحدة من اثنين من البويضات بعد حوالي 7 أيام. في أصناف غير متوافقة مع الذات التي أليل S من حبوب اللقاح هو نفسه واحد من اثنين من pistils، أنبوب حبوب اللقاح توقف عن النمو في النمط العلوي، ومنع الإخصاب (الشكل 2ج). ومع ذلك، فإن أنابيب حبوب اللقاح من أصناف متوافقة، مع أليل S مختلفة، يمكن أن تنمو من خلال نمط (الشكل 2D)، تصل إلى المبيض (الشكل 2هـ) وتخصيب واحدة من اثنين من البويضات.
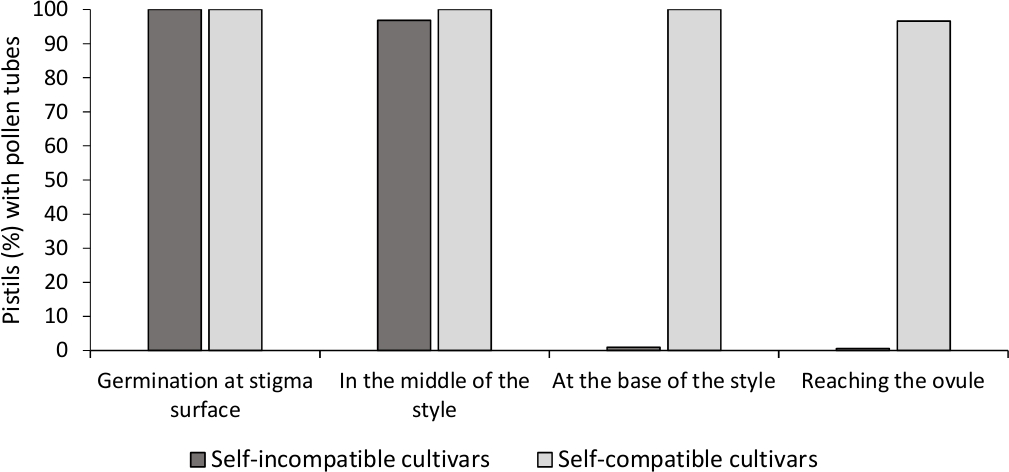
أظهر تحليل إنبات حبوب اللقاح في المختبر قابلية جيدة لحبوب اللقاح في جميع الأصناف المحللة هنا ، لأن معظم أنابيب حبوب اللقاح كانت أطول من طول حبوب اللقاح بعد 24 ساعة في الوسط الثقافي. وقد لوحظت حبوب اللقاح المُنبتة عند سطح الوصمة(الشكل 2ب)في البيستيل من جميع التلقيحات، مما يشير إلى التلقيح الكافي(الشكل 3).
لتحديد التوافق الذاتي (في) لكل صنف، لوحظ سلوك أنبوب حبوب اللقاح في التلقيح الذاتي وعبر القيام به في ظروف تسيطر عليها المختبرات تحت المجهر الفلوريسنس. تم تسجيل نمو أنبوب حبوب اللقاح على طول النمط في جميع pistils فحصها. كانت تعتبر أصناف ذاتية غير متوافقة عندما تم القبض على نمو أنبوب حبوب اللقاح على طول النمط في معظم pistils التلقيح الذاتي(الشكل 2C, الشكل 3) والنفس متوافق عندما وصلت واحدة على الأقل أنبوب حبوب اللقاح إلى قاعدة النمط في معظم pistils فحصها (الشكل 2E, الشكل 3).
وأتاحت دراسة المَلَغُس S-locusبواسطة تحليل PCR تحديد النمط الجيني S لكلصنف. أولاً، تم تحديد S-alleles بواسطة تضخيم أول S-RNase intron باستخدام التمهيدي SRc-F/SRc-R(الجدول 2). تم تحليل حجم الشظايا المضخمة بواسطة الكهرباء الشعرية(الشكل 4أ)واستخدمت لتصنيف الأنماط الجينية التي تم تحليلها في مجموعة عدم التوافق المقابلة (I.G.) (الجدول3).
وأظهرت بعض أزواج من الغيلات، مثل S1 و S7 أو S6 و S9،أحجام أجزاء مماثلة للينترون الأول. وهكذا، تم تمايز هذه الغيلة من خلال تضخيم منطقة intron الثاني من RNase مع التمهيديات برو-C2/PruC4R، SHLM1/SHLM2 SHLM3/SHLM4(الجدول 2). تم استخدام مجموعة التمهيدي PruC2/PruC4R للتمييز بين S6 و S9. S S6، تم تضخيم جزء من 1300 BP في حين لوحظ جزء من حوالي 700 bp للـ S9 allele (الشكل 4B، الجدول 3). ال التمهيديين محددة SHLM1/SHLM2 و SHLM3/SHLM4 تضخيم جزء من حوالي 650 BP في1 Sأليل و 413 bp في 7 Sأليل (الشكل 4C، الجدول 3).
ال التمهيديات AprFBC8-(F/R) التي تضخيم المناطق المتغيرة V2 وHVb من الجين SFB استخدمت للتمييز Sج و S8 alleles منذ كل من alleles تظهر تسلسل RNase متطابقة. وأظهر 8 أليل S8 PCR-جزء من حوالي 150 BP في حين أن 500 bp جزء يتوافق مع أليل Sc (الشكل 4D,الجدول 3). مرة واحدة تم تحديد S-genotype لجميع الأصناف، تم تعيين أصناف غير متوافقة ذاتيا لمجموعات عدم التوافق المقابلة على أساس Sبهم -alleles(الجدول 3).
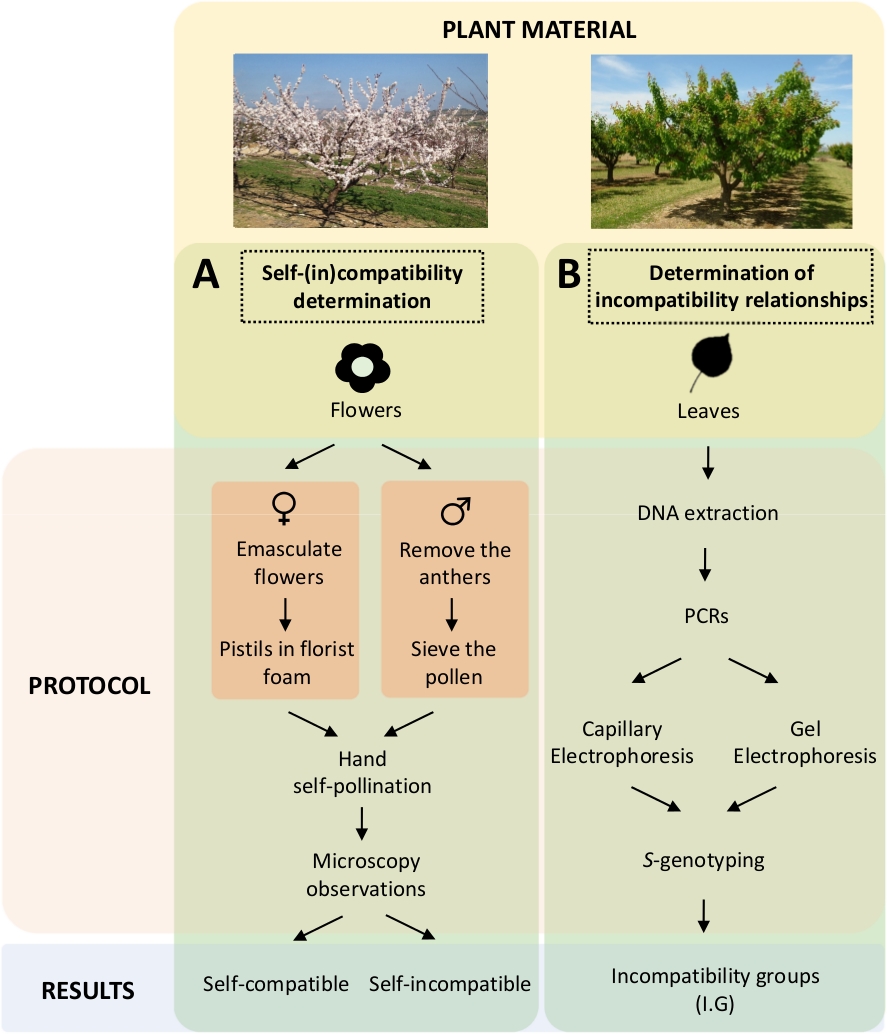
ويتطلب هذا النهج تحديد التوافق الذاتي (في) لكل صنف بواسطة التلقيحات الذاتية والتلقيحات المتبادلة الخاضعة للرقابة في المختبر(الشكل 5ألف)بالتزامن مع توصيف النمط الجيني Sبواسطة التحليل الجيني(الشكل 5B). ونتيجة لذلك، يمكن تحديد متطلبات التلقيح لكل صنف وعلاقات عدم التوافق بين أصناف المشمش.

الشكل 1- الـ 1 إعداد تجريبية لتحديد الذات (في) التوافق في المشمش.
(أ) الزهور في مرحلة البالون (السهام السوداء) في هذا المجال. (B) منخل حبوب اللقاح باستخدام شبكة غرامة. (C) Pistils وضعت على رغوة بائع الزهور في الماء. (D) تلقيح اليد من pistils بمساعدة من فرشاة الطلاء. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 2- الانبعاثات 2 من 100 التمثيل التخطيطي لعلاقات عدم التوافق gametophytic في الزهور المشمش.
(أ) في لعبة عدم التوافق الذاتي (GSI) ، على حد سواء حبوب اللقاح متوافقة وغير متوافقة تنبت على وصمة العار. حبوب اللقاح تحمل واحدة من اثنين من s-alleles من النمط الجيني الأصلي، في هذه الحالة إما S1 أو S2. إذا كان S-allele من حبوب اللقاح مباريات واحدة من اثنين من s-alleles من pistil، في هذه الحالة S1S3، يتم تثبيط نمو أنبوب حبوب اللقاح في الثلث العلوي من النمط. S (ب) إنبات حبوب اللقاح على سطح وصمة العار. (C) حبوب اللقاح أنبوب القبض في نمط تشير إلى سلوك غير متوافق. (D) حبوب اللقاح أنابيب تنمو على طول النمط. (E) أنابيب حبوب اللقاح في قاعدة النمط مما يدل على سلوك متوافق. أشرطة مقياس، 100 ميكرومتر. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 3- الانبعاثات 100-11 نتائج تمثيلية من انبات حبوب اللقاح ونمو أنبوب حبوب اللقاح من خلال نمط للأصناف متوافقة مع الذات وغير متوافقة مع الذات بعد التلقيح الذاتي.
النسبة المئوية من pistils مع حبوب اللقاح تنبت في سطح وصمة العار، مع أنابيب حبوب اللقاح في منتصف الطريق النمط، في قاعدة النمط، والوصول إلى المبيض. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 4- الانبعاثات 100 تضخيم جزء PCR باستخدام خمس مجموعات زوج التمهيدي لتحديد S-alleles.
(أ)إخراج محلل الجينات لSRc-(F/R) التمهيدية التي تبين حجم اثنين من شظايا مضخمة من المنطقة intron RNase الأولى المقابلة لS-alleles. (B) تضخيم PCR باستخدام التمهيدي PruC2/PruC4R لتحديد من 6 S6 و 99 أليل. (C) منتجات PCR التي تم الحصول عليها باستخدام التمهيديين المحددة SHLM1 و SHLM2 للتمييز بين1 أليل Sو SHLM3 و SHLM4 للتمييز مع أليل S7. (D) تضخيم PCR مع التمهيديات AprFBC8-(F/R) لتحديد Sc و S8 alleles. MI: 1 KB سلم الحمض النووي. MII: 100 BP سلم الحمض النووي. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 5- الانبعاثات 100-10 مخطط من التصميم التجريبي لتوضيح الذات وبين (في) علاقات التوافق في أصناف المشمش.
(أ)سير العمل الذاتي (في) تحديد التوافق عن طريق التلقيحات الخاضعة للرقابة في المختبر. (ب) سير عمل S-أليل تحديد بواسطة النهج الجزيئية. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
| PCR ماجستير ميكس | ظروف الدراجات الحرارية | ||||||
| مكونات | التركيز النهائي | 15 ميكرولتر التفاعل | خطوة دورة | درجه الحراره | الوقت | دورات | |
| 10x NH4 رد فعل العازلة | 10x | 1.5 ميكرولتر | التكون الأولي | 94 درجة مئوية | 3 دقائق | 1 | |
| 50 mM MgCl2 الحل | 25 mM | 1.2 ميكرولتر | Denaturing | 94 درجة مئوية | 1 دقيقة | 35 | |
| 100 mM dNTP | 2.5 مليون متر | 0.6 ميكرولتر | الصلب | 55 درجة مئوية | 1 دقيقة | ||
| التمهيدي SRc-F | 10 ميكرومتر | 0.6 ميكرولتر | ملحق | 72 درجة مئوية | 3 دقائق | ||
| التمهيدي SRc-R | 10 ميكرومتر | 0.6 ميكرولتر | التمديد النهائي | 72 درجة مئوية | 5 دقائق | 1 | |
| 500 يو طق الحمض النووي البوليميراز | 0.5 يو | 0.2 ميكرولتر | 4 درجة مئوية | عقد | |||
| H2O | 8.3 ميكرولتر | ||||||
| مكونات | التركيز النهائي | 25 ميكرولتر رد فعل | خطوة دورة | درجه الحراره | الوقت | دورات | |
| 10x PCR العازلة | 10x | 2.5 ميكرولتر | التكون الأولي | 94 درجة مئوية | 2 دقيقة | 1 | |
| 5x Q-الحل | 5x | 5 ميكرولتر | Denaturing | 94 درجة مئوية | 10 س | 10 | |
| 100 mM dNTP | 2.5 مليون متر | 0.5 ميكرولتر | الصلب | 55 درجة مئوية | 2 دقيقة | ||
| التمهيدي بروك2 | 10 ميكرومتر | 0.2 ميكرولتر | ملحق | 68 درجة مئوية | 2 دقيقة | ||
| التمهيدي C4R | 10 ميكرومتر | 0.2 ميكرولتر | Denaturing | 94 درجة مئوية | 10 س | 25 | |
| 250 يو طق DNA بوليميراز | 10 ش | 0.13 ميكرولتر | الصلب | 58 درجة مئوية | 2 دقيقة | ||
| H2O | 15.5 ميكرولتر | التمديد* | 68 درجة مئوية | 2 دقيقة | |||
| التمديد النهائي | 72 درجة مئوية | 5 دقائق | 1 | ||||
| 4 درجة مئوية | عقد | ||||||
| * مع 10 s تضاف كل دورة إلى 68 ٪ C تمديد الخطوة. | |||||||
| مكونات | التركيز النهائي | 25 ميكرولتر رد فعل | خطوة دورة | درجه الحراره | الوقت | دورات | |
| 10x PCR العازلة | 10x | 2.5 ميكرولتر | التكون الأولي | 94 درجة مئوية | 2 دقيقة | 1 | |
| 5x Q-الحل | 5x | 5 ميكرولتر | Denaturing | 94 درجة مئوية | 30 س | 35 | |
| 100 mM dNTP | 2.5 مليون متر | 0.5 ميكرولتر | الصلب | 62 درجة مئوية | 1.5 دقيقة | ||
| التمهيدي SHLM1 | 10 ميكرومتر | 0.2 ميكرولتر | ملحق | 72 درجة مئوية | 2 دقيقة | ||
| التمهيدي SHLM2 | 10 ميكرومتر | 0.2 ميكرولتر | التمديد النهائي | 72 درجة مئوية | 5 دقائق | 1 | |
| 250 يو طق DNA بوليميراز | 10 ش | 0.13 ميكرولتر | 4 درجة مئوية | عقد | |||
| H2O | 15.5 ميكرولتر | ||||||
| مكونات | التركيز النهائي | 20 ميكرولتر رد فعل | خطوة دورة | درجه الحراره | الوقت | دورات | |
| 5x PCR العازلة | 5x | 4 ميكرولتر | التكون الأولي | 98 درجة مئوية | 30 س | 1 | |
| dNTP | 2.5 مليون متر | 1.6 ميكرولتر | Denaturing | 98 درجة مئوية | 10 س | 35 | |
| التمهيدي SHLM3 | 10 ميكرومتر | 1 ميكرولتر | الصلب | 56 درجة مئوية | 30 س | ||
| التمهيدي SHLM4 | 10 ميكرومتر | 1 ميكرولتر | ملحق | 72 درجة مئوية | 1 دقيقة | ||
| 100 يو الحمض النووي بوليميراز | 5 ش | 0.2 ميكرولتر | التمديد النهائي | 72 درجة مئوية | 5 دقائق | 1 | |
| H2O | 12.4 ميكرولتر | 4 درجة مئوية | عقد | ||||
| مكونات | التركيز النهائي | 25 ميكرولتر رد فعل | خطوة دورة | درجه الحراره | الوقت | دورات | |
| 10x PCR العازلة | 10x | 2.5 ميكرولتر | التكون الأولي | 94 درجة مئوية | 2 دقيقة | 1 | |
| 100 mM dNTP | 2.5 مليون متر | 2 ميكرولتر | Denaturing | 94 درجة مئوية | 30 س | 35 | |
| التمهيدي FBC8-F | 10 ميكرومتر | 1 ميكرولتر | الصلب | 55 درجة مئوية | 1.5 دقيقة | ||
| التمهيدي FBC8-R | 10 ميكرومتر | 1 ميكرولتر | ملحق | 72 درجة مئوية | 2 دقيقة | ||
| 250 يو طق DNA بوليميراز | 10 ش | 0.125 ميكرولتر | التمديد النهائي | 72 درجة مئوية | 5 دقائق | 1 | |
| H2O | 17.4 ميكرولتر | 4 درجة مئوية | عقد | ||||
الجدول 1- الانبعاثات 1000 رد فعل وظروف ركوب الدراجات لمجموعات التمهيدي مختلفة المستخدمة في هذا البروتوكول.
| الاشعال | تسلسل | مرجع |
| SRc-F | 5'-CTCGCTTTCCTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| برو-C2 | 5'-CTTTGGCCAAGTATATATAAACC-3' | 35 |
| برو-C4R | 5'-GGATGTGGTACGATTGAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGAGAGCTGTAGGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTGTATGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCTCTCTTAG-3' | 26 |
الجدول 2- التمهيديات المستخدمة في هذا البروتوكول، تسلسل والمرجع لتوصيف S-النمط الجيني في أرمينياكا.
| أصناف | SRc-(F/ R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (BP) | AprFBC8 - (F / R) (bp) | S-نمط وراثي | مجموعة عدم التوافق (I.G) |
| وندر كوت8 | 420, 420 | 749, 1386 | S6S9 | الثامن | |||
| سحر سرير8 | 334, 420 | 749 | S2S9 | Xx | |||
| جولدسترايك8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | أنا | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| جيم - ٦ | 334, 408 | 413 | S2S7 | رابعا | |||
| كوبر كوت8 | 274, 408 | 650 | S1S3 | الثامن عشر | |||
| Apriqueen | 358, 358 | 500 | Sجج | - | |||
| بيرغيكوت8 | 334, 358 | 500 | S2Sج | - | |||
| ربيع أحمر الخدود8 | 274, 358 | 150 | S3S8 | الحادي |
الجدول 3- الانبعاثات 100 من 10 S-genotyping من أصناف المشمش مع خمسة أزواج التمهيدي المستخدمة في هذا البروتوكول وعدم التوافق تعيين المجموعة. مختلف البوليميراز سلسلة التفاعل أحجام المنتجات S-alleles تضخيم باستخدام SRc -(F / R)، PruC2/PruC4R، SHLM1 / SHLM2، SHLM3/SHLM4، و AprFBC8-(F/ R) يتم عرض التمهيديين في الجدول.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
تقليديا، معظم أصناف المشمش التجارية الأوروبية متوافقة مع36. ومع ذلك، فإن استخدام أصناف أمريكا الشمالية غير المتوافقة ذاتيا كوالدين في برامج التربية في العقود الماضية أدى إلى الإفراج عن عدد متزايد من أصناف جديدة غير متوافقة ذاتيا مع متطلبات التلقيح غير معروف7،8،37. وهكذا ، فإن تحديد الذات وبين (في) علاقات التوافق في أصناف المشمش هو على نحو متزايد أهمية. وتزداد حدة هذه في تلك المناطق حيث تقشعر لها الأبدان في فصل الشتاء في تناقص، منذ ارتفاع الاختلافات من سنة إلى سنة في وقت المزهرة تمنع من مصادفة في ازدهار الأصناف وحبوب اللقاح في كثير من الحالات، وخاصة في الأصناف مع متطلبات تقشعر لها الأبدان عالية38. وقد كانت المنهجية الموصوفة هنا، والجمع بين التلقيح اليدوي، والمجهر والتحليلات الجينية مفيدة جدا لتحديد التوافق الذاتي (في) كل صنف، وإنشاء أصنافها المحتملة في مجال التلقيح.
ويمكن تحديد متطلبات التلقيح من خلال تجارب التحكم الميداني في ظروف البساتين11،39. ومع ذلك، فإن المعرض لعوامل خارجية بما في ذلك الظروف الجوية السلبية يمكن أن يسبب فشل التلقيح10، مما قد يؤدي إلى تشخيص خاطئ من عدم التوافق الذاتي. تسمح المنهجية الموصوفة هنا بتقييم التوافق الذاتي (في) بشكل أكثر دقة بواسطة عمليات المجهر للزهور الملقحة يدوياً في ظروف يتم التحكم فيها مختبرياً، مع تجنب التأثير البيئي. وعلاوة على ذلك، يسمح هذا النهج تحليل عدد أكبر من أصناف سنويا، لأن هناك حاجة سوى عدد قليل من الزهور بدلا من العديد من الأشجار الكبار لكل أصناف التي هي مطلوبة في التجارب الميدانية40.
يمكن إقامة علاقات عدم التوافق بين التلقيح اليدوي والمجهر14. ومع ذلك ، لا يمكن إجراء التلقيحات إلا لفترة قصيرة خلال موسم الإزهار في الربيع ، وهناك حاجة إلى أشجار البالغين بالقرب من المختبر ، لأن عمر الزهور التي يتم جمعها قصير للغاية. وهكذا، فإن عدد علاقات عدم التوافق التي يمكن تحليلها عن طريق التلقيحات اليدوية التي تسيطر عليها في كل موسم منخفض جدا. وقد مكن توصيف الجينات المشفرة من قبل S-locus تطوير أساليب PCR القائم على لS-أليلgenotyping18,41. هذا النهج يسرع S-أليه تحديد لأنه لا يتطلب الزهور، ويمكن إجراء التجارب مع أي الأنسجة النباتية42. هذا يمدّد الفترة أثناء أيّ مادة نباتيّة, عادة يترك شابّة, يستطيع كنت جمعت43. وعلاوة على ذلك، يمكن أن يترك lyophilized أو المجمدة، بحيث يمكن أن يتم التحليل في أي وقت من السنة، على عكس التلقيحات التي يمكن القيام به فقط على الزهور الطازجة خلال موسم المزهرة44. فائدة إضافية هي أنه يمكن جمع الأوراق من الأشجار الصغيرة حتى قبل دخول عصر الإزهار ، مما يسهل جمع العينات والحصول المبكر على النتائج45.
التحليل الجيني يسمح بتمايز أفضل من الأليل عدم التوافق الذاتي لأنه يوفر نتائج دقيقة لأحجام جزء تضخيم21,46. حتى الآن، تم تحديد ثلاثة وثلاثين S-alleles في المشمش12،18،20،21،22،23،24، والذي سمح لإنشاء 36 مجموعات عدم التوافق على أساس S -النمط الجيني8،9،17،25،26،27. ومن ناحية أخرى، فإن العيب في هذه المنهجية هو أن أليل مختلفة في نفس حجم النطاق أو الطفرات يمكن أن تحدد خطأ مثل أليل نفسه. وهكذا، Sج و S8 أليل متطابقة لسلسلة RNase ولكن تم العثور على الإدراج 358-bp في الجين SFB من Sc19. وبالمثل، فإن منطقة intron الأولى من الاليليس S1 و S7 متطابقة ويمكن اشتفرافها باستخدام التمهيدي SRc-F/SRc-R. بالإضافة إلى ذلك، تم العثور Sعلى العديد من الـ 6 و S528 أو S20 و S55و S7 و S6 13 (EF062341) و S4617،لأن بعض هذه الاليات تم تسلسلها جزئيًا أو بسبب الفشل أثناء تضخيم PCR، وبالتالي، هناك حاجة إلى مزيد من العمل لتمييزها بشكل صحيح.7
تحليل PCR وتسلسل S-RNase كافية لإقامة علاقات عدم التوافق من خلال تحديد S-alleles وتخصيص أصناف في مجموعة عدم التوافق المقابلة الخاصة بهم8،17،26،27. S ومع ذلك ، فإن هذه المنهجية لديها حدود منع تحديد الذات (في) التوافق لأصناف مشمش معين. وقد ارتبطت التوافق الذاتي (SC) إلى خاصة S-alleles في الأنواع الأخرى من القرنية 47، كما اللوز (Sو)48،49 أو الحلو الكرز (S4')50،51. ومع ذلك، في المشمش، يمكن أن يتم تحديد الأليل Sc، الذي ارتبط بـ SC21،خطأً على أنه S8،وأليل غير متوافق ذاتيًا19،22، والطفرات المحتملة غير المرتبطة بملوط S، حيث تم تحديد M-locus12و52. في الآونةالأخيرة، تم ا ا ا ا ا ا ا ا ا ا ا ا ا ا ا ا ا استخدام SSR علامات12. ولذلك، فإن التحديد الجيني لـ SC للنمط الجيني المشمش يحتاج إلى مزيد من البحث، ومن أجل تجنب الأخطاء بسبب عوامل غير مرتبطة بـ S، في هذا العمل تم تحديد توصيف التوافق الذاتي (في) أيضًا من خلال الظواهر سلوك أنابيب حبوب اللقاح من خلال pi الزهور ذاتية التلقيح.
المنهجية الموصوفة هنا التي تجمع بين تحديد التوافق الذاتي (في) التوافق عن طريق التلقيح اليدوي في ظروف المختبر مع الملاحظة اللاحقة لسلوك أنابيب حبوب اللقاح في مبيل التلقيح الذاتي الخاضع للرقابة تحت المجهر الفلوري وتحديد النمط الجيني Sبواسطة تحليل PCR يسمح بتحديد متطلبات التلقيح من أصناف المشمش. وهذا يوفر معلومات قيمة للمزارعين والمربين، لأنه يسمح بإقامة علاقات عدم التوافق بين الأصناف لاختيار الملقحات المناسبة في تصميم البساتين الجديدة وكذلك لاختيار الآباء المناسبين لتصميم الصلبان الجديدة في برامج تربية المشمش.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
ليس لدى أصحاب البلاغ ما يكشفون عنه.
Acknowledgments
تم تمويل هذا البحث من قبل وزير دي سينسيا، Innovación y Universidades-Fund European Regional Development Fund، والاتحاد الأوروبي (AGL2016-77267-R، و AGL2015-74071-JIN)؛ المعهد الوطني للاستثمارات و تيكنولوجيا أغريا y أليمنتاريا (RFP2015-00015-00, RTA2017-00003-00)؛ Gobierno de Aragón -European Social Fund, European Union (Grupo Consolidado A12_17R), Fundación Biodiversidad and Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}