

Summary
הטרוגניות תוך-גידולית היא תכונה אינהרנטית של גידולים, כולל גליומות. פיתחנו פרוטוקול פשוט ויעיל המשתמש בשילוב של חוצצים וצנטריפוגה הדרגתית כדי לבודד גרעינים בודדים מרקמות גליומה קפואות טריות למחקרי רצף גרעין יחיד ו- ATAC.
Abstract
גליומות מפוזרות למבוגרים מציגות הטרוגניות בין-גידולית. עד לאחרונה, רוב מאמצי הפרופיל המולקולרי בקנה מידה גדול התמקדו בגישות בתפזורת שהובילו לסיווג המולקולרי של גידולים במוח. במהלך חמש השנים האחרונות, גישות רצף תאים בודדים הדגישו כמה תכונות חשובות של gliomas. רוב המחקרים הללו השתמשו בדגימות גידול במוח טרי כדי לבודד תאים בודדים באמצעות ציטומטריית זרימה או שיטות הפרדה מבוססות נוגדנים. במבט קדימה, השימוש בדגימות רקמות קפואות טריות מביו-בנקים יספק גמישות רבה יותר ליישומי תאים בודדים. יתר על כן, ככל שהשדה של התא הבודד מתקדם, האתגר הבא יהיה ליצור נתונים מרובי אומיקה מתא בודד או מאותה הכנה לדוגמה כדי לפענח טוב יותר את מורכבות הגידול. לכן, פרוטוקולים פשוטים וגמישים המאפשרים יצירת נתונים עבור שיטות שונות כגון רצף RNA חד-גרעין (snRNA-seq) וגרעין יחיד Assay עבור Chromatin נגיש Transposase עם רצף תפוקה גבוהה (snATAC-seq) יהיה חשוב עבור התחום.
ההתקדמות האחרונה בתחום התא הבודד בשילוב עם מכשירים מיקרופלואידיים נגישים כגון פלטפורמת הגנומיה 10x הקלו על יישומי תאים בודדים במעבדות מחקר. כדי לחקור את ההטרוגניות של גידול במוח, פיתחנו פרוטוקול משופר לבידוד גרעינים בודדים מגליומות קפואות טריות. פרוטוקול זה ממזג פרוטוקולים קיימים של תא בודד ומשלב שלב הומוגניזציה ואחריו סינון וצנטריפוגה הדרגתית בתיווך מאגר. הדגימות המתקבלות הן השעיות גרעין יחיד טהור שניתן להשתמש בהם כדי ליצור ביטוי גן גרעין יחיד נתוני נגישות כרומטין מאותה הכנת גרעין.
Introduction
גליומות מפוזרות בדרגה נמוכה יותר (LGG), הגידול המוחי העיקרי הנפוץ ביותר במבוגרים, הן גידולים חודרניים המתעוררים לעתים קרובות בחצי הכדור המוחי. LGGs מפגינים הטרוגניות בין-גידולית, המונעת לא רק על ידי אוכלוסיית הגידול אלא גם על ידי התאים הלא ממאירים המעורבים באופן מורכב בהתפתחות הגידול ובהתקדמות1,2,3,4,5.
בעשור האחרון, יש מפולת של נתונים גנומיים שנאספו בתחום גליומות. נתונים אלה מגיעים בעיקר ממחקרי ריצוף גידול בתפזורת ותרמו רבות לאפיון המולקולרי, והסיווג הנוכחי של גידולים במוח5,6,7,8,9,10,11. עם זאת, למרות שמחקרים אלה חשפו את הנוף המולקולרי הרחב הקשור לגלומות, עדיין קיים חוסר התקדמות מאכזב לגבי התערבות טיפולית. אחד המכשולים לטיפול בהתנגדות בגידולים במוח הוא הטרוגניות תוך-גידולית. כדי לטפל בבעיה זו, מחקרים שונים התמקדו בהטרוגניות הגנומית, התמלולית, הפרוטאומית והאפיגנטית הקיימת בתוך גידול ברמת תא בודד12,13,14,15,16,17.
למרות שחלו התפתחויות טכנולוגיות יוצאות דופן בתחום התא הבודד בשנים האחרונות, אחד הגורמים המגבילים העיקריים הוא הזמינות של דגימות טריות הדרושות כדי לבודד את התאים ולבצע את הניסויים האלה. כדי להתגבר על מגבלה זו, היו כמה ניסיונות מוצלחים לבצע ניסיונות, כגון snRNA-seq ו snATAC-seq מרקמות קפואות, באמצעות גרעינים ולא תאים18,19. רוב השיטות הללו מסתמכות על מיון תאים המופעלים על-ידי פלואורסצנטיות (FACS) או על אסטרטגיות סינון. הן לגישות של תא בודד והן לגרעין יחיד יש את נקודות החוזק והחסרונות שלהם. גישות תא בודד לשמור על תמלילים מיטוכונדריאליים, אשר למרות עשוי להיות אינפורמטיבי, יכול גם להפחית את כיסוי שעתוק בשל השפע הגבוה שלהם. גישות בידוד גרעינים יחיד לחסל אחוז גבוה של שבר המיטוכונדריה, ובכך לאפשר כיסוי מעמיק יותר של תמלילים גרעיניים20.
ישנן פלטפורמות מסחריות שונות ששימשו בשנים האחרונות כדי לאבדק נתוני גנומיקה של תאים בודדים, כולל RNA-seq ו- ATAC-seq. אחת הפלטפורמות הבולטות ביותר היא פלטפורמת כרום גנומיקה 10x לביטוי גנים של תא בודד ופרופיל ATAC של תא יחיד. כאשר הפלטפורמה פועלת בעזרת תאים מיקרופלואידיים, כל פסולת או אגרגטים יכולים לסתום את המערכת המובילה לאובדן נתונים, ריאגנטים ודגימות קליניות יקרות ערך. לכן, ההצלחה של מחקרי תא יחיד תלויה במידה רבה בבידוד מדויק של תאים בודדים / גרעינים.
הפרוטוקול שנדגים כאן הוא שילוב שונה במקצת של פרוטוקולי DroNc-seq ו- Omni-ATAC-seq, ומשתמש בגישה דומה למחקרים אחרונים המשתמשים ב- snRNA-seq כדי להבין הפרעות נוירולוגיות וסוגי תאים עצביים במוחהאנושי 18,19,21,22,23,24. הפרוטוקול משתמש בשילוב של דיסוציאציה אנזימטית/מכנית של דגימות קפואות ואחריו סינון וצנטריפוגה הדרגתית ומאפשר בידוד מהיר ומדויק של גרעינים בודדים מרקמות גליומה קפואות טריות. השתמשנו בהצלחה בפרוטוקול זה כדי ליצור נתוני snRNA-seq ו- snATAC-seq מאותה הכנת גרעין מדגימות גידול במוח.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
דגימות גליומה קפואות טריות התקבלו מהמרכז הלאומי למחלות גידול (NCT) - בנק רקמה בהיידלברג, גרמניה. השימוש בחומר מטופלים אושר על ידי הוועדה לביקורת מוסדית בפקולטה לרפואה בהיידלברג, והושגו הסכמה מדעת מכל המטופלים הכלולים במחקר.
1. הכנה ניסיונית
- בצע את כל השלבים על קרח או ב 4 °C (7 °F).
- צינורות טרום צמרמורת, כלים, סכיני גילוח, Douncers ועלים עד 4 °C (70 °F).
- הכן את כל המאגרים מראש. מאגרים אלה יציבים בטמפרטורת החדר. סינון סטרילי מומלץ, במיוחד עבור סוכרוז. ההכנות למניה משתנות מקורס ואח '19. ראה טבלאות 1-7.
- הסר דגימות מחנקן נוזלי או אחסון מקפיא -80 °C ולשמור על קרח יבש עד שלב 2.1.
2. ניתוח רקמות וניתוק
- מעבירים דגימת רקמות קפואות טריות (10-60 מ"ג) לצלחת פטרי מצוננת מראש. טחון / לקצוץ רקמה קפואה טרייה עם סכין גילוח לחתיכות קטנות על קרח.
- הוסף 500 μL של גרעין מצונן תמוגה חוצץ צינור 1.5 מ"ל צונן מראש. מניחים את חתיכות הרקמה בצינור 1.5 מ"ל המכיל את מאגר תמוגתהגרעינים, ומעביריםל- Douncer ( שולחן החומרים).
הערה: ישנם שני עלים הומוגניזר Dounce: "רופף" או "A" עלה להפחתת מדגם ראשוני "הדוק" או "B" עלה עבור הומוגניזציה מדגם מלא. - Dounce חתיכות הרקמה עם העלי "רופף" במשך כ 20 משיכות, עד החיכוך מופחת. אם קיים פסולת, המדגם עשוי להיות פינה מראש על ידי סינון עם מסננת 100 מ"מ.
- Dounce עם עלה "הדוק" עבור 20 משיכות כדי להשיג הומוגניזציה רקמות מלאה.
- מעבירים את ההומוגנאט (כ-500 μL) לצינור 2 מ"ל צונן מראש. הוסף 1 מ"ל של מאגר תמה מצוננת. מערבבים בעדינות ודגרה על קרח במשך 5 דקות. מערבבים בעדינות עם קצה פיפטה רחב-נשא וחוזרים על הפעולה 1-2 פעמים במהלך הדגירה.
- לסנן את ההומוגנית כולה באמצעות רשת מסננת 30 מיקרומטר, לאסוף לתוך צינור פלקון 15 מ"ל ולהעביר בחזרה לתוך צינור חדש 2 מ"ל מקורר מראש. מסננת אחת בדרך כלל מספיקה לכל ההומוגנית.
- בדוק תחת מיקרוסקופ אור כדי לאמת את הסרת פסולת גדולה ואת שלמות הממברנה הגרעינית. גרעינים צריכים להיות עגולים ואין לעוות את הממברנה הגרעינית. אם קיימים פסולת, ניתן לסנן מחדש גרעינים.
- צנטריפוגה הגרעינים ב 500 x g במשך 5 דקות ב 4 °C (5 °F) על צנטריפוגה העליון ספסל. הסר את supernatant, משאיר מאחור ~ 50 μL עם גלולה המכילה את הגרעינים. בעדינות resuspend הכדור עוד 1 מ"ל של חוצץ תמוגה גרעינים ודגרה במשך 5 דקות על קרח.
- צנטריפוגות הגרעינים ב 500 x גרם במשך 5 דקות ב 4°C. הסר את supernatant ללא גלולה מטרידה, להוסיף 500 מ"ל של 1x חוצץ הומוגניזציה (HB) (טבלה 4) ודגרה במשך 5 דקות ללא שימוש חוזר. לאחר מכן, resuspend הגרעינים עוד 1.0 מ"ל של 1x HB.
- צנטריפוגה הגרעינים ב 500 x גרם במשך 5 דקות ב 4 °C (5 °F). הסר את supernatant בעדינות resuspend הגרעינים ב 200 μL של 1x HB לתוך צינור חדש 2.0 מ"ל.
3. צנטריפוגה הדרגתית
- הוסף 200 μL של 50% תמיסה יודיקסנול (טבלה 5) כדי לתת ריכוז סופי של 25% יודixanol. מערבבים היטב 10 פעמים עם פיפטה שנקבעה על 300 מ"ל.
- הוסף 300 μL של 29% תמיסה יודיקסנול (טבלה 6) תחת תערובת 25%. השתמש בקצה דק P1000 כדי למנוע תערובת של השכבות.
- הוסף 300 μl של 35% תמיסה יודיקסנול (טבלה 7) תחת תערובת 29%. השתמש בקצה דק P1000 כדי למנוע תערובת של השכבות.
אזהרה: שלב זה דורש הסרה הדרגתית של קצה הפיפטה במהלך ההצינורות כדי למנוע תזוזה מוגזמת של נפח. - מניחים את הדגימות בצנטריפוגת דלי מתנדנדת, מסתובבים במשך 20 דקות ב 3,500 x g ב 4 ° C עם הבלם כבוי.
- הסר בעדינות את הדגימות מבלי לרעוד והתבונן תחת אור. רצועה לבנה ברורה של 95% גרעינים טהורים צריכה להיות גלויה בין השכבה השנייה והשלישית (איור 1).
4. בידוד גרעינים
- שאפו את השכבות העליונות עד לרצועת הגרעין הלבנה באינטרפאזה של 29%-35%.
- לאסוף את רצועת גרעין בנפח 200 מ"ל, להעביר צינור טרי לסנן עם מסנן 20 מיקרומטר(טבלת חומרים).
הערה: הגרעינים אינם צריכים להיות respended לפני סינון. - בדוק תחת מיקרוסקופ אור כדי לאמת את הסרת פסולת גדולה ואת שלמות הממברנה הגרעינית. גרעינים צריכים להיות עגולים ואין לעוות את הממברנה הגרעינית.
- ספירת גרעינים באמצעות כתמים כחולים טריפאן על hemocytometer ו nuclei aliquot עבור snRNA-seq / snATAC-seq.
5. גרעין יחיד RNA ו- ATAC seq
- מעבדים מיד את הגרעינים באמצעות ביטוי הגן של התא הבודד וערכות ריאגנט ATAC של תא בודד(טבלת החומרים).
הערה: ריכוזי דגימת הגרעינים עבור מערכת הגנומיקה 10x הם 1500-3000 גרעינים לכל μL עבור snRNA-seq, ו 3500-7000 גרעינים למ"ר μL עבור snATAC-seq. הגרעינים יכולים להיות מדוללים באמצעות PBS 1x. - רצף את הספריות המתקבלות במתקן הליבה הגנומי.
- בצע ניתוח בקרת איכות של הנתונים. גרעינים כלולים לניתוח נוסף אם הם מכילים מזהה מולקולרי ייחודי (UMI) >1000, מספר הגנים >500 ואחוז מהגנים המיטוכונדריאליים <5%, והם בתוך ממוצע + שלוש סטיות תקן של UMIs וגנים.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
גנומיקה של גרעין יחיד היא שדה מתפתח עם נתונים ופרוטוקולים מוגבלים. גורם קריטי המשפיע על התוצאה של התקפות גרעינים יחידים הוא בידוד של גרעינים טהורים ושלמים. שילבנו שני פרוטוקולים שפורסמו (פרוטוקולי DroNc-seq ו- Omni-ATAC-seq) כדי לבודד גרעינים איכותיים וטהורים מבלוקי רקמת גליומה קפואים טריים תוך זמן קצר יחסית ובכך לשמור על יציבות התמלילים (איור 1).
השימוש בשלבי סינון שונים יחד עם צנטריפוגה הדרגתית באמצעות שיפוע יודיקסנול/סוכרוז מאפשר בידוד של גרעינים טהורים כאשר רוב הפסולת מושלכת(איור 2). ניתן להשתמש באותה הכנת גרעינים מבודדת הן עבור snRNA-seq והן עבור snATAC-seq. חשוב לציין, מאחר שהגרעין המשמש הוא מאותה דגימה, ניתן להטמיע את הנתונים שנוצרו באמצעות חבילות כגון סרה כדי ליצור אשכולות ולספק פרספקטיבה מרובת אומיקות25 (איור 3).
כדי לקבוע אם הפרוטוקול דומה ערכות נתונים snRNA-seq שפורסמו, השווינו נתונים שהושגו באמצעות ההליך עם ארבעה מחקרים snRNA-seq זמין לציבור הקשורים למערכת העצבים המרכזית (CNS): סליפר ואח'20,אגם ואח'26, ג'יקל ואח '24 וחביב ואח'18. כדי להשוות בין מדדי בקרת האיכות, הורדנו את מטריצות הספירה הבאות מ- Omnibus ביטוי הגנים (GEO): GSE104525 (ערכת נתונים חביב, 2017), GSE97930 (ערכת נתונים של אגם, 2018), GSE118257 (ערכת נתונים של ג'יקל, 2019) ו- GSE140819 (ערכת נתונים של סליפר, 2020). עבור ערכת הנתונים של האגם, מטריצת ספירה גולמית נפוצה נוצרה על ידי מיזוג מטריצות בודדות לחצי הכדור המוחי, קליפת המוח הקדמית וקליפת המוח החזותית. עבור ערכת הנתונים של סליפר, נבחרה ספירה גולמית עבור המדגם HTAPP-443-SMP-5491 (גליומה ילדים ברמה גבוהה).
כדי לבצע השוואה משוחדת, כל הדגימות למעט ערכת הנתונים של ג'יקל עובדו באמצעות פרוטוקול סטנדרטי משותף. ראשית, השתמשנו בחבילה הסטטיסטית של סרה R כדי ליצור אובייקט סרה של כל מטריצה גולמית27. לאחר מכן נעשה שימוש בשני שלבים: 1) להסרת טיפות פוטנציאליות, נעשה שימוש בקיצוצים הבאים - גרעינים המכילים פחות מ-1,000 UMI, פחות מ-500 גנים או יותר מ-5% מה-RNA המיטוכונדריאלי לא נכללו בניתוח ו-2) כדי לא לכלול חריגים, גרעינים שנפלו מחוץ לממוצע בתוספת שלוש סטיות תקן להפצת UMIs וגנים הוסרו. עבור ערכת הנתונים של Jakel, לא היתה אפשרות לבצע שלב שני זה, מכיוון שערכת הנתונים הזמינה לציבור טופלה מראש עם שלב בקרת איכות פחות מחמיר.
כדי להשוות את ההתפלגות של UMIs וגנים בין דגימות, מיזבנו את כל ערכות הנתונים ודמיינו את ההתפלגות של מספר UMIs וגנים באמצעות חלקתכינור (איור 4). תוצאה זו הצביעה על כך שהשיטה דומה לפרוטוקול snRNA-seq העדכני ביותר המתואר ב- Slyper et al.20.
הפרוטוקול המומחשה כאן עוסק בדגימות גליומה, אך ניתן ליישם את אותה גישה באופן ריאלי עבור גידולים ורקמות שאינם CNS. עם זאת, זה ידרוש אופטימיזציה של הרכבי מאגר תמותה זמני דגירה.
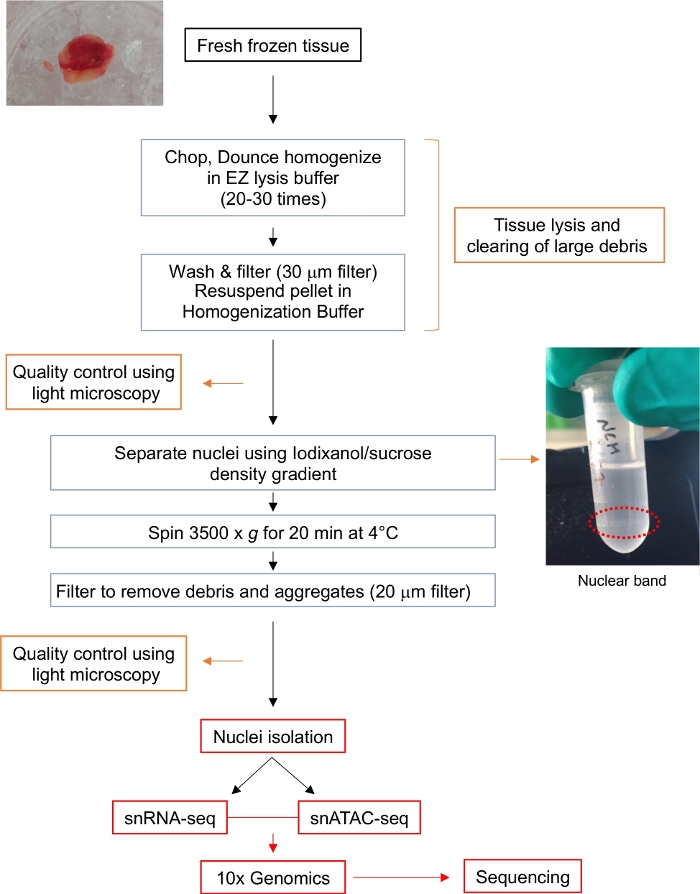
איור 1: תרשים זרימה לבידוד גרעינים. תרשים הזרימה מספק מבט קצר על השלבים המעורבים בבידוד של גרעינים בודדים מרקמת גליומה קפואה טרייה. תמונות מייצגות עבור דגימת הגידול ואת הלהקה הגרעינית לאחר שיפוע Iodixanol / סוכרוז (מוקף בקו מנוקד אדום) מוצגים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
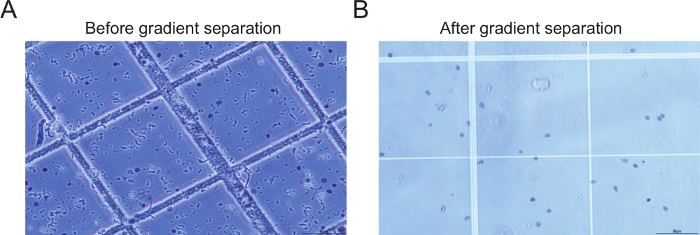
איור 2: דוגמאות לגרעין לפני ואחרי צנטריפוגה הדרגתית. (A)התמונה של המדגם לפני ביצוע צנטריפוגה הדרגתית וסינון מראה כמויות גדולות של פסולת (B) התמונה של המדגם לאחר צנטריפוגה הדרגתית וסינון שלב מראה גרעינים שלמים עם כמות מינימלית של פסולת. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 3: דוגמה לנתונים מוטבעים מ-snRNA-seq ו-snATAC-seq באמצעות אותה הכנת גרעין. החבילה הסטטיסטית Seurat R שימשה לשילוב נתוני snRNA-seq ו- snATAC-seq27. (A)תמונה מוטבעת משותפת של snRNA-seq ו- snATAC-seq data (B) אשכולות המיוצרים על ידי הטבעה משותפת של נתוני snRNA-seq ו- snATAC-seq. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.

איור 4: פרמטרי בקרת איכות של ערכות נתונים שונות במוח האנושי snRNA-seq המציגים את המספר הבודד של UMI (A) ומספר הגנים (B) לכל גרעינים. מספר הגרעינים שעברו מסנני איכות היה כדלקמן: 3527 גרעינים ממערת הנתונים של סלייפר, 14636 גרעינים ממערת הנתונים של נריאנן שנוצרו באמצעות הפרוטוקול המתואר, 7369 גרעינים ממערת הנתונים של ג'יקל, 16494 גרעינים ממערת הנתונים של האגם ו-4652 גרעינים ממערת הנתונים של חביב. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
| 6x חיץ הומוגניזציה יציבה תערובת מאסטר | ||
| מגיב | קונק סופי. | כרך עבור 100 (מ"ל) |
| 1 מ' קcl2 | 30 מ"ר | 3.0 |
| 1 מ"ג (Ac)2 | 18 מ"ר | 1.8 |
| 1 M טריס pH 7.8 | 60 מ"ר | 6.0 |
| ח2O | 89.2 | |
| שמור בטמפרטורת החדר, הימנע מחשיפה ישירה לאור | ||
טבלה 1: הכנת 6x חוצץ הומוגניזציה יציב תערובת מאסטר.
| 1 M סוכרוז |
| 34.23 גרם סוכרוז |
| להתמוסס ב 78.5 מ"ל של מים |
| מלא עד 100 מ"ל במים |
טבלה 2: הכנת 1 M סוכרוז.
| 6x חיץ הומוגניזציה פתרון לא יציב (650 מ"ל לדגימה) | ||
| מגיב | קונק סופי. | Vol לכל דגימה (μL) |
| 6x חוצץ הומוגניזציה יציב | 6x | 648.84 |
| 100 mM PMSF (פנילמתילסולפוניל פלואוריד) | 0.1 מ"מ | 1.08 |
| 14.3 מ' β-מרקפטונול | 1 מ"מ | 0.08 |
טבלה 3: הכנת 6x חיץ הומוגניזציה פתרון לא יציב (650 μL לדגימה).
| 1x פתרון לא יציב של מאגר הומוגניזציה (2 מ"ל לדגימה) | ||
| מגיב | קונק סופי. | Vol לכל דגימה (μL) |
| 6x חוצץ הומוגניזציה לא יציב | 1x | 333.33 |
| 1 M סוכרוז | 320 מ"ר | 640.00 |
| 50 מ"מ EDTA | 0,1 מ"מ | 4.00 |
| 10% NP40 | 0.1% | 20.00 |
| ח2O | 1006.27 | |
טבלה 4: הכנת פתרון לא יציב של מאגר הומוגניזציה 1x (2 מ"ל לדגימה).
| 50% תמיסה יודיקסנול (200 μL לדגימה) | ||
| מגיב | קונק סופי. | Vol לכל דגימה (μL) |
| 6x חוצץ הומוגניזציה לא יציב | 1x | 66.67 |
| 60% תמיסה יודיקסנול | 50% | 333.33 |
טבלה 5: הכנת 50% תמיסה יודיקסנול (200 μl לדגימה).
| 29% תמיסה יודיקסנול (300 μL לדגימה) | ||
| מגיב | קונק סופי. | Vol לכל דגימה (μL) |
| 6x חוצץ הומוגניזציה לא יציב | 1x | 100 |
| 1 M סוכרוז | 160 מ"ר | 96 |
| 60% תמיסה יודיקסנול | 29% | 290 |
| ח2O | 114 | |
טבלה 6: הכנת 29% תמיסה יודיקסנול (300 μl לדגימה).
| 35% תמיסה יודיקסנול (300 μL לדגימה) | ||
| מגיב | קונק סופי. | Vol לכל דגימה (μL) |
| 6x חוצץ הומוגניזציה לא יציב | 1x | 100 |
| 1 M סוכרוז | 160 מ"ר | 96 |
| 60% תמיסה יודיקסנול | 35% | 350 |
| ח2O | 54 | |
טבלה 7: הכנת פתרון יודיקסנול 35% (300 μl לדגימה).
Subscription Required. Please recommend JoVE to your librarian.
Discussion
תחום ההטרוגניות התוך-גידולית נמצא בשלב מרגש, עם בדיקות ופלטפורמות חדשניות שפותחו כדי לאתגר ולהרחיב את הידע הקיים. הטרוגניות תוך-גידולית היא גורם מכריע התורם להתקדמות המחלה ולעמידות בפני שיטות הטיפול הנוכחיות בגליומות28. מחקרים אחרונים על גידולים במוח התמקדו בהיבט חשוב זה באמצעות ניתוחים תעתיקיים ואפיגנומיים של תאים בודדים כדי לאפיין טוב יותר את ההטרוגניות התאית בתוך אותו גידול29,30,31,32. אחד צווארי הבקבוק הנוכחיים עם בדיקות תאים בודדים בגידולים במוח, וגידולים מוצקים אחרים, הוא הזמינות של דגימות קליניות טריות. כדי להתגבר על בעיה זו, מחקרים שונים הראו כי באמצעות גרעינים מרקמות קפואות טריות יכול להיות חלופה דגימות טריות והוא יכול לשמש בהצלחה כדי לחקור הטרוגניות הסלולר18,33.
כאן, אנו משפרים את שיטות הבידוד הגרעיניות הבודדות הקודמות במונחים של פשטות, אורך, שפע ואיכות גרעינים. גישה זו מביאה עוד יותר את היתרון של יצירת פרופיל snRNA-seq ו snATAC-seq מאותה הכנת גרעינים, המאפשר מחקרים multi-omics. בהליך זה, שינינו פרוטוקולים קיימים על ידי הוספת תהליך נוסף של הפרדה הדרגתית באמצעות מדיום שאינו יוני מבוסס יודיקסנול. גישה זו מאפשרת בידוד של גרעינים טהורים ללא צורך במיון FACS, ואיכות הגרעינים מוערכת תחת מיקרוסקופ אור. השעיה באיכות טובה של גרעינים לא צריכה להיות גושים, פסולת מינימלית, גרעינים שלמים. למרות נדיר, גושים של גרעינים יכול להתרחש במהלך שלב הבידוד, אשר ניתן לפתור על ידי צנרת אור באמצעות טיפים פיפטה P200 רחב נשא או על ידי מאמץ את הגרעינים באמצעות מסנן 20 מ"מ. הגורם הקריטי ביותר בגנומיקה של גרעין יחיד הוא היכולת להשיג גרעינים יחידים טהורים ושלמים. נוכחות של פסולת או אגרגטים יכולה להוביל לבלוק של התאים המיקרופלואידיים בתוך פלטפורמות התא הבודד, ובכך להוביל לנתונים באיכות נמוכה או לכישלון אפשרי של הניסוי. מטרת השימוש במסננים בשלבים שונים ולאחריהם התבוננות תחת מיקרוסקופ אור היא למנוע התרחשויות כאלה.
כמות ואיכות החומר ההתחלתי (למשל, גידול קפוא טרי) הם גם שיקולים חשובים. השתמשנו בהצלחה בפרוטוקול המתואר כאן עם בלוקים רקמת גידול במוח הנע בין 10 מ"ג ל 60 מ"ג. עבור דגימות אלה, 500 μL של גרעין תמוגה חוצץ (שלב 1) מספיק כדי להשיג גרעינים באיכות טובה מספיק עבור רצף גרעין יחיד. בלוקים גידול גדול משמעותית המכילים יותר מ 60-70% תוכן הגידול ניתן לחתוך לחתיכות קטנות יותר, חתיכות ניתן גם מיד להציב בחזרה לתוך חנקן נוזלי או -80 °C אחסון או מחולק צינורות לבידוד גרעינים יחיד מעורב לאחר שלב תמוגה הושלמה. בנוסף, איכות הרקמה ותכולת הגידול צריכה להיות מוערכת על ידי הכתמת Hematoxylin ו- Eosin (H&E) ואחריה אישור של פתולוג.
הגורמים החשובים הבאים שיש לקחת בחשבון הם הטמפרטורה וזמן העיבוד. אחסון רקמות לא תקין, טיפול בדגימות ופרוטוקולים ארוכים עלולים להשפיע לרעה על איכות נתוני הגנומיה הסופיים. הדגימות צריכות להישמר על קרח, והזמן לעיבוד מדגם צריך להיות מהיר כדי למנוע השפלה של גרעינים. העיבוד הממוצע עבור מדגם אחד עם פרוטוקול זה הוא 45-60 דקות, ולוקח כשעתיים לעבד ארבע דגימות. אחסון רקמות לא תקין יכול להשפיע גם על בידוד גרעינים ואיכות נתונים. הערכנו אם משך זמן האחסון ב- -80 °C (80 °F) השפיע על איכות הנתונים שנוצרו באמצעות הפרוטוקול המתואר. השוואת תוצאות הרצף שהתקבלו מהדגימות הקפואות בין השנים 1 ל-14 שנים (2006-2019) לא הראתה הבדלים באיכות הנתונים ביחס לזמן האחסון. לכן, דגימות רקמות קפואות קפואות כראוי יכולות לשמש למחקרים רטרוספקטיביים באמצעות פרוטוקול זה.
ישנן מספר מגבלות עם פרוטוקולי גרעין יחיד. לדוגמה, לא ניתן למיין סוגי תאים שונים בהתבסס על סמני פני השטח של התא, ותעתיקים ציטופלסמיים אינם ניתנים לזיהוי. כמו כן, אנו להשמיט את התמלילים המיטוכונדריאליים ומכאן לאבד על מידע ביולוגי מסוים הקשורים חילוף החומרים של הגידול. יתר על כן, סוגי רקמות שונים עשויים לדרוש מאגרי תמותה שונים ובכך דורשים אופטימיזציה של הפרוטוקול עם כל התקנה ניסיונית חדשה.
בסך הכל, הפרוטוקול המתואר כאן הוא פשוט, מקטין את זמן העיבוד לדוגמה, ומניב גרעינים באיכות גבוהה. היעדר שלב מיון מפחית את הלחץ על הגרעינים ומבטל את הצורך במיון תאים. היעדר RNA מיטוכונדריאלי מגביר את עומק הרצף של תמלילים גרעיניים. חשוב לציין, בדומה לפרוטוקולי בידוד גרעין יחידים אחרים, פרוטוקול משופר זה מאפשר שימוש בדגימות קפואות בארכיון לניתוח רטרוספקטיבי. יישמנו בהצלחה זרימת עבודה זו כדי לבצע snRNA-seq ו snATAC-seq באמצעות אותה הכנת גרעין מגידולים במוח ללא צורך לשנות מאגרים. לכן, גישה זו מאפשרת מחקרים מרובי omics מאותה דגימה.
לסיכום, שיטת הבידוד של גרעין יחיד המתוארת כאן היא טכניקה יעילה, מדויקת ומהירה ביותר אשר ניתן ליישם כדי לבצע מחקרי ריצוף גרעין יחיד.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
למחברים אין מה לחשוף.
Acknowledgments
אנו מודים למעבדה הפתוחה של תא יחיד (scOpenLab) במרכז הסרטן הגרמני (DKFZ) על דיונים מועילים. מחקר זה נתמך על ידי הסיוע הגרמני לסרטן, מקס-Eder תוכנית מענק מספר 70111964 (S.T.).
Materials
| Name | Company | Catalog Number | Comments |
| 2-Mercaptoethanol | Sigma | M6250 | |
| CaCl2 | Sigma | 21115-100ML | |
| Dounce Homogenizer | Active motif | 40401 | |
| EDTA (0.5 M) | Thermo Scientific | R1021 | |
| Falcon 15 mL Conical Centrifuge Tubes | Fisher Scientific | 352096 | |
| Iodixanol (aka Optiprep) | Stem cell technologies | 07820 | |
| MACs Smart Strainers (30 µm) | Miltenyi Biotec | 130-098-458 | |
| MACS SmartStrainers (100 µm) | Miltenyi Biotec | 130-098-463 | |
| Mg(Ac)2 | Sigma | 63052-100ML | |
| NP-40 | Abcam | ab142227 | |
| Nuclei Isolation Kit: Nuclei EZ Prep | Sigma | NUC101-1KT | |
| Phenylmethanesulfonyl fluoride (PMSF) | Sigma | P7626 | |
| Pre-Separation Filters (20 µm) | Miltenyi Biotec | 130-101-812 | |
| Safe lock tubes 1.5 mL | Eppendorf | 0030120086 | |
| Safe lock tubes 2.0 mL | Eppendorf | 0030120094 | |
| Single Cell ATAC | 10x Genomics | ||
| Single Cell Gene Expression | 10x Genomics | ||
| Sucrose | Sigma | S0389 | |
| Wide Bore pipette tips (1000 µL) | Themo Fisher Scientific | 2079GPK | |
| Wide Bore pipette tips (200 µL) | Themo Fisher Scientific | 2069GPK |
References
- Huse, J. T., Holland, E. C. Targeting brain cancer: advances in the molecular pathology of malignant glioma and medulloblastoma. Nature Reviews Cancer. 10 (5), 319-331 (2010).
- Kreso, A., Dick, J. E. Evolution of the cancer stem cell model. Cell Stem Cell. 14 (3), 275-291 (2014).
- Filbin, M. G., Suva, M. L. Gliomas Genomics and Epigenomics: Arriving at the Start and Knowing It for the First Time. Annual Review of Pathology. 11, 497-521 (2016).
- Ferris, S. P., Hofmann, J. W., Solomon, D. A., Perry, A. Characterization of gliomas: from morphology to molecules. Virchows Archive. 471 (2), 257-269 (2017).
- Louis, D. N., et al. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: a summary. Acta Neuropathologica. 131 (6), 803-820 (2016).
- Eckel-Passow, J. E., et al. Glioma Groups Based on 1p/19q, IDH, and TERT Promoter Mutations in Tumors. The New England Journal of Medicine. 372 (26), 2499-2508 (2015).
- Suzuki, H., et al. Mutational landscape and clonal architecture in grade II and III gliomas. Nature Genetics. 47 (5), 458-468 (2015).
- Cancer Genome Atlas Research. Comprehensive, Integrative Genomic Analysis of Diffuse Lower-Grade Gliomas. The New England Journal of Medicine. 372 (26), 2481-2498 (2015).
- Noushmehr, H., et al. Identification of a CpG island methylator phenotype that defines a distinct subgroup of glioma. Cancer Cell. 17 (5), 510-522 (2010).
- Yan, H., et al. IDH1 and IDH2 mutations in gliomas. The New England Journal of Medicine. 360 (8), 765-773 (2009).
- Yan, H., Bigner, D. D., Velculescu, V., Parsons, D. W. Mutant metabolic enzymes are at the origin of gliomas. Cancer Research. 69 (24), 9157-9159 (2009).
- Gawad, C., Koh, W., Quake, S. R. Single-cell genome sequencing: current state of the science. Nature Reviews Genetics. 17 (3), 175-188 (2016).
- Tanay, A., Regev, A. Scaling single-cell genomics from phenomenology to mechanism. Nature. 541 (7637), 331-338 (2017).
- Wu, M., Singh, A. K. Microfluidic Flow Cytometry for Single-Cell Protein Analysis. Methods in Molecular Biology. 1346, 69-83 (2015).
- Schwartzman, O., Tanay, A. Single-cell epigenomics: techniques and emerging applications. Nature Reviews Genetics. 16 (12), 716-726 (2015).
- Macaulay, I. C., Ponting, C. P., Voet, T. Single-Cell Multiomics: Multiple Measurements from Single Cells. Trends in Genetics. 33 (2), 155-168 (2017).
- Buenrostro, J. D., et al. Single-cell chromatin accessibility reveals principles of regulatory variation. Nature. 523 (7561), 486-490 (2015).
- Habib, N., et al. Massively parallel single-nucleus RNA-seq with DroNc-seq. Nature Methods. 14 (10), 955-958 (2017).
- Corces, M. R., et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nature Methods. 14 (10), 959-962 (2017).
- Slyper, M., et al. A single-cell and single-nucleus RNA-Seq toolbox for fresh and frozen human tumors. Nature Medicine. 26 (5), 792-802 (2020).
- Mathys, H., et al. Single-cell transcriptomic analysis of Alzheimer's disease. Nature. 570 (7761), 332-337 (2019).
- Krishnaswami, S. R., et al. Using single nuclei for RNA-seq to capture the transcriptome of postmortem neurons. Nature Protocols. 11 (3), 499-524 (2016).
- Al-Dalahmah, O., et al. Single-nucleus RNA-seq identifies Huntington disease astrocyte states. Acta Neuropathologica Communications. 8 (1), 19 (2020).
- Jakel, S., et al. Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature. 566 (7745), 543-547 (2019).
- Butler, A., Hoffman, P., Smibert, P., Papalexi, E., Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nature Biotechnology. , (2018).
- Lake, B. B., et al. Integrative single-cell analysis of transcriptional and epigenetic states in the human adult brain. Nature Biotechnology. 36 (1), 70-80 (2018).
- Stuart, T., et al.
- Bready, D., Placantonakis, D. G.
- Venteicher, A. S., et al. Decoupling genetics, lineages, and microenvironment in IDH-mutant gliomas by single-cell RNA-seq. Science. 355 (6332), (2017).
- Tirosh, I., et al. Single-cell RNA-seq supports a developmental hierarchy in human oligodendroglioma. Nature. 539 (7628), 309-313 (2016).
- Weng, Q., et al. Single-Cell Transcriptomics Uncovers Glial Progenitor Diversity and Cell Fate Determinants during Development and Gliomagenesis. Cell Stem Cell. 24 (5), 707-723 (2019).
- Neftel, C., et al. An Integrative Model of Cellular States, Plasticity, and Genetics for Glioblastoma. Cell. 178 (4), 835-849 (2019).
- Al-Ali, R., et al. Single-nucleus chromatin accessibility reveals intratumoral epigenetic heterogeneity in IDH1 mutant gliomas. Acta Neuropathologica Communications. 7 (1), 201 (2019).
