1. Microscope set up and imaging preparation
- Our imaging set up consists of a Leica MZ16 fluorescent stereomicroscope with a fast shutter and a cooled CCD camera controlled by Metamorph software.
- Prepare a thermostatically controlled heating pad and adjust output to 32.5°C to maintain the animal’s body temperature during and following surgery.
- Warm sterile Ringer’s solution or artificial cerebrospinal fluid (ACSF) to 32.5°C in advance for irrigation of spinal cord during surgery.
- Anesthetize the animal with an intraperitoneal injection of xylazine (8 mg/kg) and ketamine (120 mg/kg) cocktail.
- Shave the upper back with small animal clippers and spread one small drop of hair removal lotion over the shaved area with cotton-tipped swabs. Minutes later, remove the applied lotion using 70% ethanol soaked gauze sponges.
2. Laminectomy and surgical exposure of the L5 dorsal root
- Place the animal on a heated (32.5° C) pad and disinfect the skin with 70% ethanol soaked swabs.
- Under bright field illumination on the stereomicroscope, perform a midline incision (2- to 3-cm) in the skin of the back. If necessary, use cotton-tipped swabs to stop bleeding.
- Reflect the spinal musculature to expose the underlying lumbar vertebrae.
- Expose the L3-S1 spinal segments by right-sided hemi-laminectomy using small rongeurs. A partial laminectomy exposing 4-6 segments of the lumbar and sacral spinal cord is created by removing the right dorsal portion of the L5 vertebrae at the level of the iliac crest of the hip (location of L5 DRG) rostrally to the L2 vertebrae (2 vertebrae caudal to the last rib). Perfuse the cavity with warm sterile Ringer’s solution.
- Position the animal on a support cushion (rolled cotton gauze) to flatten the vertebral column. Use retraction hooks to widen the exposed area.
- At this point (approximately 30 minutes after the first i.p. injection of anesthetic, a supplement (0.5X) should be injected subcutaneously in order to keep the animal fully anesthetized. Alternatively, gas anesthesia (2-4% isoflurane in 0.5L/min oxygen) can be used for repeated anesthetization during imaging sessions longer than 1 hour.
3. Rhizotomy/Dorsal root crush
- Switch to fluorescence excitation to visualize YFP labeled (+) axons.
- Using the tip of a Sub-Q (26ga.) needle, perform a small incision in the dura overlying the L5 dorsal root (DR). Perfuse repeatedly with Ringer’s solution and clean gently with cotton-tipped swabs.
- Identify the site to be crushed and insert one side of a fine forceps (Dumont #5) subdurally.
- Close the forceps gently but firmly, holding the medial portion of the L5 root for 10 seconds, and then gently release the forceps.
- Wash repeatedly with physiological solution and clean gently with cotton-tipped swabs.
4. Image acquisition and post-op procedures
- Obtain multiple images of the whole exposed area including the crush site and the DREZ before and right after the crush at both low and high magnification.
- Images are acquired either as single snapshots or as multiple streams of 10 to 20 frames acquired within 30- to 40-ms exposure time. In-focus images are then selected, and an overview montage is created later using Photoshop.
- To minimize scar formation, tightly apply a piece of thin synthetic matrix membrane (Biobrane), followed by artificial dura, on top of the exposed spinal cord. Be sure to cut pieces to fit precisely in the exposed spinal cord window so that they are adherent to the spinal cord.
- Close the musculature with sterile 5-0 sutures and close the midline incision with wound clips.
- Inject Ringer’s solution (0.3- to 0.5-mL subcutaneously), and administer Buprenorphine as postoperative analgesia (0.05 mg/kg) subcutaneously every 12 h for 2 days.
- Keep the animal on a heating pad (34°-35° C) until recovered.
5. Repeated imaging
- Anesthetize the animal and remove the wound clips and sutures.
- Remove artificial dura and thin synthetic matrix membrane patches and keep them in a sterile tube containing Ringer’s solution for later reuse.
- Gently remove any accumulated connective tissue scar with the bent tip of a Sub-Q needle and fine forceps. Perfuse frequently with warm Ringer’s solution.
- Re-expose the operative field, including crush site and DREZ, relocate YFP+ axons imaged in previous sessions, and repeat the procedures described in Section 4.
6. Representative results:
We have observed that, whereas they did not regenerate after a transection injury, almost all YFP+ axons grew through the site of injury by 3 days after crush (Figure 1)11. Typically, the next day after crush, we observed dying-back degeneration of proximal stump axons and fragmentation/degeneration of the same axons distal to the crush, which confirmed that axons had been appropriately damaged (e.g., Figure 1; Day 3 and 5). Several additional criteria are applied to unambiguously distinguish regenerating axons from axons that were spared or had recovered from the injury. These included the following: (1) regenerating axons show an expansion of the non-fluorescent portion of the YFP+ axon at the crush site due to proximal and distal degeneration (in contrast to narrowing of the unlabeled gap due to fluorescent cytoplasm refilling the crush site if axons survived the injury); (2) regenerating axons are much thinner, less brightly fluorescent, and more undulating than axons that survived the injury; (3) regenerating neurites are thinner and more dimly fluorescent than the degenerating fluorescent fragments of axons through which they extended; (4) in contrast to surviving or spared axons, regenerating axons stop at the DREZ; and (5) in contrast to surviving or spared axons, regenerating axons do not exhibit nodes of Ranvier. Figure 1 shows four superficial YFP+ axons immediately after crush (A1; colored arrows). Three days after the crush, all four axons extend a single neurite that grows through the crush site (A2). Five days after crush, neurites remain stable and there is no additional growth from these or other proximal axons (A3).
The regenerating neurites that crossed the crush site elongated through much thicker and brighter fluorescent fragments of a degenerating axon (i.e., endoneurial tubes), and arrive at the DREZ as early as 4 days after the crush (about 3mm/2 days)11. Repeated imaging of these axons and their tips every two or three days for two more weeks (Figure 2) revealed that they did not grow forward or retract, but remained immobile. The only noticeable change was swelling of the tips and shafts of some axons. These observations therefore demonstrate surprisingly quick and chronic immobilization of the regenerating axons at the DREZ.
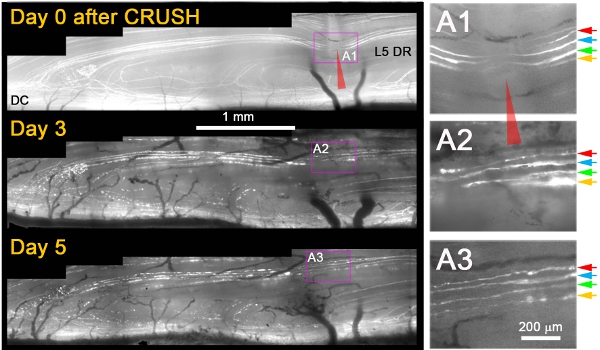
Figure 1: Repeated imaging of L5 DR YFP+ axons at the site of dorsal root crush over 5 days. The medial portion of the L5 root was crushed (red arrowhead) and imaged on Days 0, 3, and 5 after the crush. The area of the crush is magnified in the right panels (A1-A3).
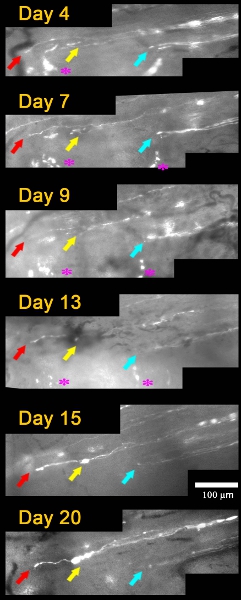
Figure 2: Repeated imaging of axons arrived at the DREZ over 20 days after L5 root crush. Day 4, three axons (colored arrows) arrived at the DREZ. The tips of these axons remain in the same location and have a similar appearance in subsequent imaging sessions on Days 7, 9, 13, 15, and 20. Positions of an axon tip relative to other axon tips and landmarks (asterisks) were used to determine axon motility between imaging sessions.
