We have shown previously that in the corneal organ cultures, the differences in the expression of diabetic markers (e.g., basement membrane proteins and integrin α3β1) and wound healing between the normal and diabetic corneas are preserved. This culture system was subjected to the gene therapy aimed at normalizing the levels of diabetes-altered markers, c-met, MMP-10, and cathepsin F.
When the whole corneal epithelium was transduced with the AV-cmet, or AV-shRNA to MMP-10 or cathepsin F (separately or in combination), the epithelial wound healing was completed significantly faster (P<0.03 by paired Student t test compared to AV-vector) than in the AV-vector-transduced fellow corneas from the same donors16,17. The AV-cmet reduced the healing time by half, which did not significantly differ from healing of the normal corneas (Figure 1). The AV-shM10 and AV-shCF had smaller effect. However, a combination of the AV-cmet with the AV-shM10 and AV-shCF (Combo) produced the strongest effect completely normalizing the epithelial healing times (Figure 1). This reduction in healing time was accompanied by the normalized patterns of diabetic (basement membrane and integrin) and stem cell markers in the diabetic corneas8,16,17.
The epithelial stem cells are the major wound healing participants4. Therefore, we examined whether limbal gene therapy targeting the stem cell compartment would be beneficial for the diabetic corneas. To this end, only the limbal parts of the diabetic corneas were transduced by the AV-cmet or Combo and large, 8.5 mm wounds were created. The AV-cmet transduced corneas healed significantly faster (P <0.001 by paired Student t test) than the AV-vector transduced fellow corneas (Figure 2). The Combo treatment resulted in the same outcome18. Gene therapy increased the expression of diabetes-suppressed markers, such as integrin α3β1 and basement membrane component nidogen-1 (Figure 3, left columns, Combo). Importantly, as in the whole corneal transduction8,17, the limbal gene therapy with either the AV-cmet or Combo led to a markedly increased expression of putative LESC markers including K15 and ΔNp63α, compared to the AV-vector transduced corneas (Figure 3, right columns, both c-met and Combo). These data supported the role of stem cell normalization in the accelerated wound healing upon gene therapy of diabetic corneas and show the feasibility of our approach to the treatment of diabetic corneal disease.
We next started to examine the gene therapy effects on cultured LESC-enriched limbal epithelial cells. The cultures of normal and diabetic limbal epithelial cells were established and stained for putative LESC markers. Many cells stained positive (Figure 4). The staining intensity in the diabetic cultures was consistently weaker than in the normal cultures (K15 shown as an example in Figure 4), which was very similar to the situation in the diabetic vs. normal ex vivo corneas8. These experiments validated the obtained cell cultures as suitable for the gene therapy.
Preliminary experiments with a telomerase-immortalized corneal epithelial cell line (see23) showed that the cells were easily transduced by the available AV constructs (data not shown here). However, the primary limbal cultures proved to be much more sensitive to the viral load and/or the transduction reagent, necessitating optimization of the transduction protocol. The cultured limbal cell transduction by the AV-GFP (Figure 5) depended upon the multiplicity of infection (MOI; range 30-300 pfu/cell) resulting in a fairly high level of transduction at high MOI (GFP expression in >80% cells). However, the higher MOI were also toxic, causing cell death or severely impaired cell migration into the scratch wounds after 3-4 days of transduction (Figure 6). At the lower MOI (10-30 pfu/cell), the AV transduction produced less or no cytotoxic effect, but resulted in a decreased number of GFP-expressing cells (5-15%).
We next attempted to boost the transduction efficiency without compromising cell viability by using transduction-enhancing reagents that generally facilitate the AV binding to the cell surface. They showed different efficiency improvements in limbal epithelial cell transduction by the AV at MOI 10-30. Both polycations, poly-L-lysine, and polybrene, were non-toxic and the most effective resulting in 3-4 fold increase in the number of GFP-positive cells (Figure 7, top row). ibiBoost was also effective but slightly toxic, and ViraDuctin was ineffective and rather toxic (Figure 7, bottom row). Therefore, polycations offer safe and efficient way to boost gene of interest expression upon the AV transduction and should be used for the gene expression and wound healing experiments.
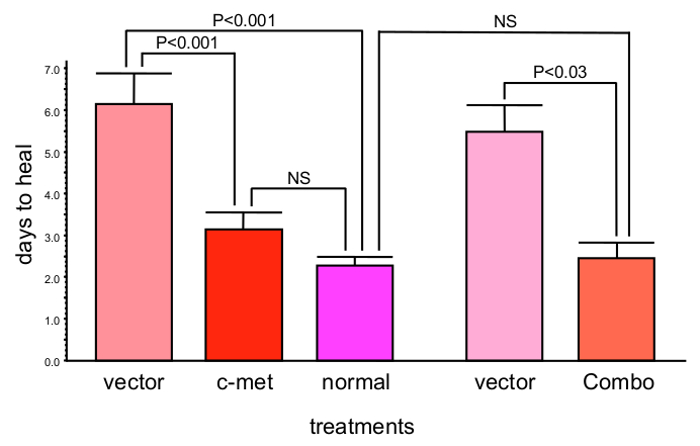
Figure 1: c-Met gene transduction leads to significantly decreased corneal epithelial wound healing time (mean ± SEM). Moreover, combined gene therapy (Combo) with AV harboring c-met gene and shRNAs to MMP-10 and cathepsin F genes completely normalizes epithelial wound healing time. Significance (p values) was established by paired Student t test in comparison with respective vector treatments. Bars represent the standard error of mean. Please click here to view a larger version of this figure.
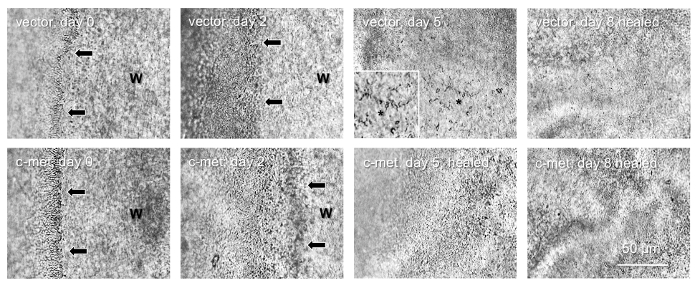
Figure 2: Healing of 8.5 mm epithelial wounds in a pair of diabetic corneas. Top row, vector-transduced cornea; healing is complete in 8 days. Bottom row, AV-cmet transduced fellow cornea; healing is complete in 5 days. Arrows show wound (W) edge. *, non-healed part (also shown at high magnification as inset), with the spider-shaped apoptosed stromal keratocytes. They are visible until the epithelial layer grows on top of them. Bar = 50 μm. Phase contrast. Please click here to view a larger version of this figure.
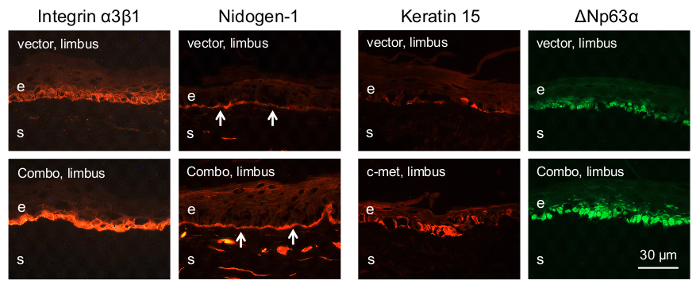
Figure 3: Immunostaining of diabetic corneal sections for various markers after limbal gene therapy. Both AV-cmet and Combo treatments result in markedly increased limbal staining for diabetic markers (integrin α3β1 and nidogen-1) and putative stem cell markers (K15 and ΔNp63α). Arrows on nidogen-1 panels mark corneal epithelial basement membrane. Bar = 30 μm. e, epithelium; s, stroma. Please click here to view a larger version of this figure.
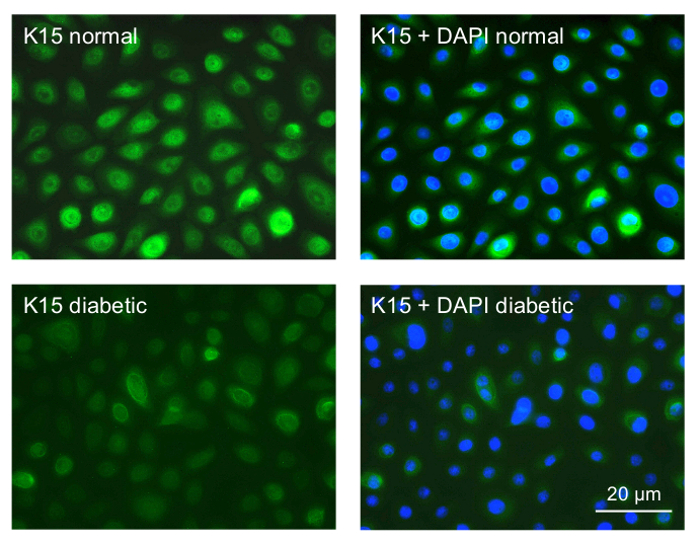
Figure 4: Immunocytochemical staining of primary limbal epithelial cultures for a putative LESC marker K15. Top row, normal culture; most cells show strong staining for K15. Bottom row, diabetic culture; most cells only show weak staining. The right panels are presented with DAPI nuclear counterstaining. Normal and diabetic cells were photographed with the same exposure time. Bar = 20 µm. Please click here to view a larger version of this figure.
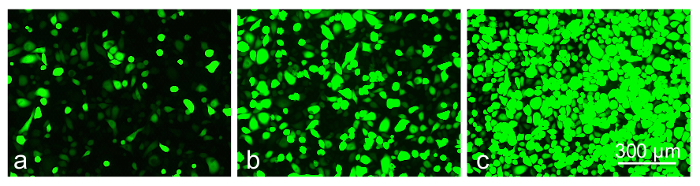
Figure 5: The level of GFP-expression in transduced limbal epithelial cells depends on MOI of AV-GFP: (a) 30 pfu/cell; (b) 120 pfu/cell; (c) 300 pfu/cell at 3 days of transduction. Photographs of live cells are shown. Bar = 300 µm. Please click here to view a larger version of this figure.
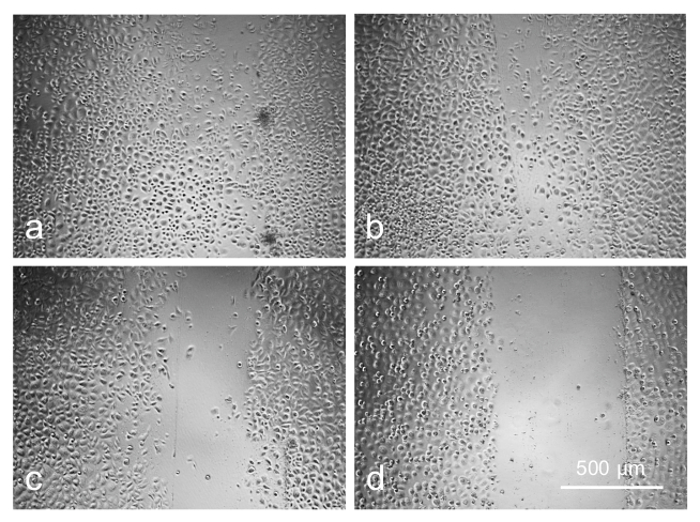
Figure 6: Cell migration is adversely affected by increasing concentration of AV as revealed by scratch wound test: (a) control; (b) 20 pfu/cell; (c) 80 pfu/cell; (d) 120 pfu/cell. Photographs of live cells are shown. Bar = 500 µm. Please click here to view a larger version of this figure.
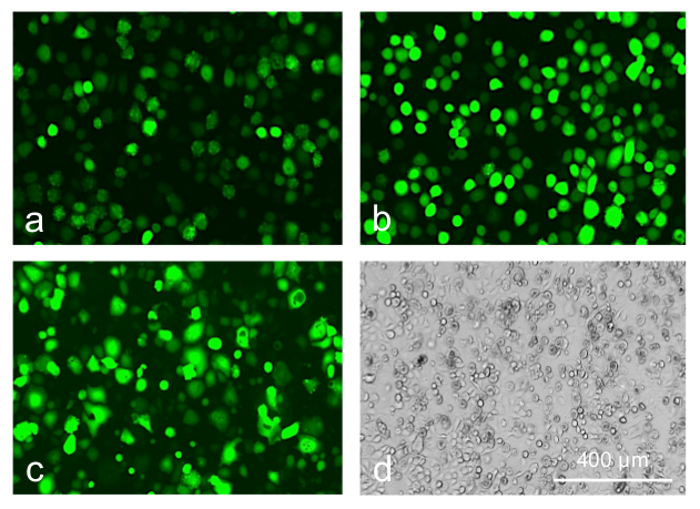
Figure 7: Limbal epithelial cells transduced with AV-GFP at 30 pfu/cell. (a) control; (b) poly-L-lysine; (c, d) transduction reagent; 5 days of transduction. Transduction reagent caused a significant cytotoxic effect leading to cell rounding and death (d, phase contrast). Photographs of live cells are shown. Bar = 400 µm. Please click here to view a larger version of this figure.
