This protocol describes two different methods to induce autophagy in mouse tissues by aerobic exercise: a total of 90 min of forced exercise on a multi-lane treadmill proceeded by two days of acclimation; or two weeks of voluntary exercise on a running wheel used by single-housed mice. In each exercise protocol, we can measure the autophagy flux by fluorescence microscopy and western blot analysis in various organs.
We used a transgenic mouse line expressing GFP-tagged LC3 as a reporter system to monitor autophagy by exercise1. Upon autophagy induction, LC3 translocates from the cytosol to the autophagosome in punctate structures. After sectioning, formation of GFP-LC3 puncta can be directly visualized by fluorescence microscopy (Figure 1A). Alternatively, autophagosome structures can also be immunostained by an LC3 antibody for imaging. Either 90 min of treadmill exercise or 2 weeks of voluntary running increased the number of GFP-LC3 puncta in both skeletal muscle (vastus lateralis) and cerebral cortex, compared to the resting condition (Figure 1B). It should be noted that the frontal cortex region has been the major region in the brain where autophagy is clearly induced by either method so far. Exercise also induced the conversion of LC3 from the cytosolic form (LC3-I) to the lipid-conjugated form (LC3-II), which can be detected by western blot analysis (Figure 1C).
Exercise-induced increment of autophagosomes (represented by LC3-II and GFP-LC3 puncta) is due to an elevated autophagic flux, rather than a block in autophagosome degradation, assessed by the use of inhibitors of lysosomal degradation, such as bafilomycin A1 or chloroquine. Here we measured the degradation of the autophagic cargo receptor p62 as an example. Exercise (90 min of treadmill) caused a higher degradation of p62 in skeletal muscle than the resting condition, which was rescued by injection with chloroquine prior to exercise (Figure 2). The similar results are also observed with voluntary exercise by running wheels. Thus, aerobic exercise by treadmill or running wheels induces autophagy in vivo, measured by the steady-state level of LC3 and the degradation of p62.
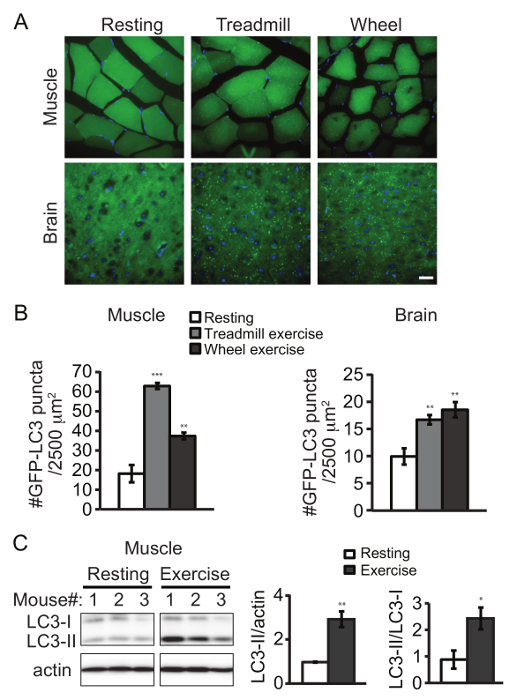
Figure 1. Aerobic Exercise Induces Autophagy in Mouse Tissues. Representative images (A) and quantification (B) of GFP-LC3 puncta in skeletal muscle (vastus lateralis) and brain (frontal cortex) of GFP-LC3 transgenic mice under the control condition (resting), after 90 min of forced exercise (treadmill) or after 2 weeks of voluntary exercise (wheel). Results represent mean ± s.e.m of 10 pictures per mouse. N = 5 mice. The following emission wavelengths were used: GFP – 525 nm; DAPI – 490 nm. (C) Western blot detection of LC3 in skeletal muscle (vastus lateralis) from rested and exercised (by treadmill) mice. Quantification data represent the level of LC3-II normalized to actin (left) and the ratio of LC3-II to LC3-I (right). N = 3 mice. Statistic is comparing each value to the control sample. Results represent mean ± s.e.m. *, P < 0.05; **, P < 0.01; ***, P < 0.001, t-test. Scale bar, 25 µm. Please click here to view a larger version of this figure.
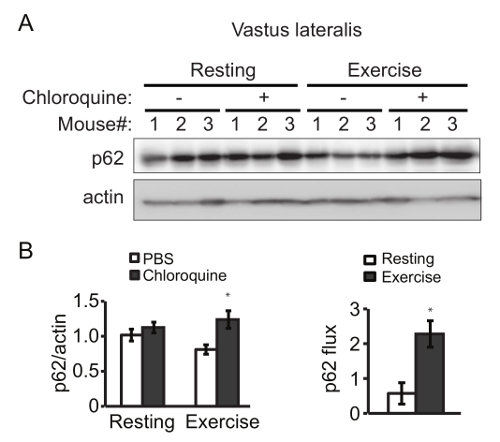
Figure 2. Exercise Increases Autophagy Flux in Mouse Skeletal Muscle. (A) Western blot detection of p62 in vastus lateralis from rested and exercised mice in the presence or absence of the lysosomal inhibitors chloroquine. (B) (Left) Quantification analysis of p62 normalized to the corresponding actin band. (Right) The p62 flux is determined by subtracting the normalized densitometric value of PBS-treated p62 from that of chloroquine-treated p62. Results represent mean ± s.e.m. N = 3 mice. *, P < 0.05, t-test. Please click here to view a larger version of this figure.
