A typical result of a hematoxylin and eosin (H&E) staining of a tibialis anterior muscle before and after cardiotoxin mediated injury is presented in Figure 4A,B. In the control muscles, the architecture of the muscle is intact as seen by the localization of the nuclei in the periphery of the myofibers and the lack of accumulation of mononucleated cells in the interstitial space (Figure 4A). 7 days after cardiotoxin mediated injury new myofibers are formed marked by centrally located nuclei (Figure 4B). Furthermore, an accumulation of mononucleated cells can be observed, consisting mostly of satellite cells but also non-myogenic cells like immune cells. The whole muscle should be injured characterized by the central location of all myonuclei and the accumulation of mononucleated cells on the whole muscle section.
To characterize the progression and success of the regeneration process, several immunofluorescent staining using markers of regeneration can be performed. The number of satellite cells can be evaluated by staining for Pax7, the canonical marker for satellite cells (Figure 4C,D). Three days after injury the number of satellite cells increases (Figure 4D), satellite cells are not located under the basal lamina anymore. To further analyze the regeneration process, newly formed myofibers can be stained with antibodies directed to developmental myosin (Figure 4E). Newly formed myofibers display centrally located nuclei and a strong expression of developmental myosin. As the myofibers mature, expression of developmental myosin decreases, the diameter of the myofiber increases while nuclei are migrating to the periphery of the myofibers.
The influence of a single gene on the process of regeneration can be investigated using transgenic mice or as shown here by injection of a self-delivering siRNA. Fluorescently labeled self-delivering scrambled control siRNA was injected into the regenerating tibialis anterior muscle at day 3 after injury, the time point when the proliferation of satellite cells peaks. Two days after injection of the siRNA satellite cells were analyzed for the presence of the fluorescently labeled siRNA (Figure 4F). We analyzed how many satellite cells had taken up the fluorescently labeled siRNA two days after injection into the regenerating muscle (Figure 5). About 75% of all satellite cells in the regenerating muscle were positive for the fluorescently labeled siRNA. Furthermore, we determined that about 74% of all regenerating myofibers were positive for the fluorescently labeled siRNA suggesting that either 74% of the regenerating myofibers had taken up the siRNA or that siRNA-positive satellite cells either had fused with each other to form new myofibers or that fusion with already existing regenerating myofibers occurred (Figure 5). This suggests that satellite cells take up the self-delivering siRNA and that the siRNA persists in the satellite cells for at least two days.
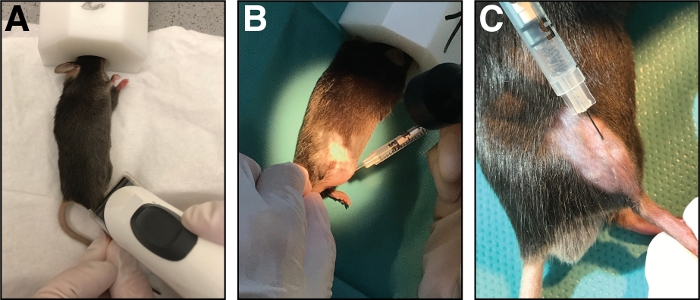
Figure 1: Injection of cardiotoxin into the tibialis anterior muscle. (A) Mice are anesthetized by inhalation of isoflurane and the lower limb is shaved. (B) A 29 G needle is injected into the tibialis anterior muscle (C) and moved along the tibia bone during injection of the cardiotoxin solution. Please click here to view a larger version of this figure.
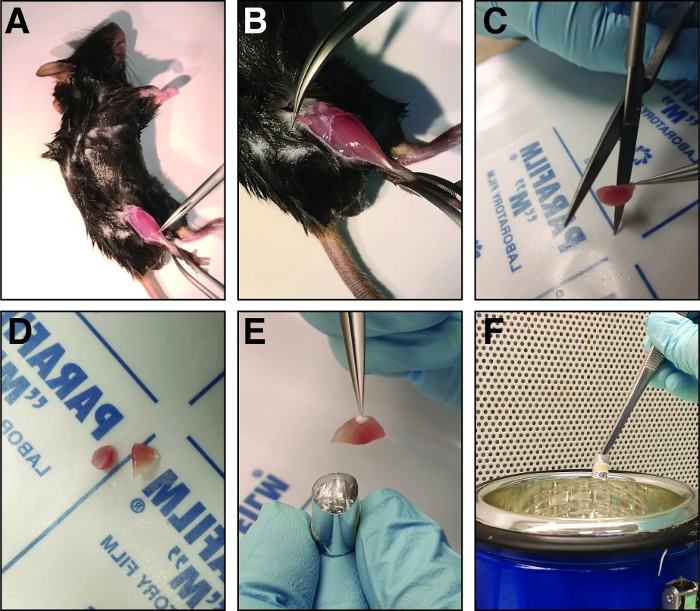
Figure 2: Steps involved in dissection and freezing of the injured tibialis anterior muscle. (A) The hind limb muscles are exposed (B) and the fascia surrounding the tibialis anterior muscle is ripped to expose the muscle. (C) After removing the muscle, it is cut at the mid belly region into two halves of similar size using sharp straight scissors (D). (E) The halves of the dissected muscle are embedded in an aluminum foil mold filled with freezing solution and frozen in liquid nitrogen (F). Please click here to view a larger version of this figure.

Figure 3: Equipment and steps involved in cryosectioning of tibialis anterior muscles. (A) The chamber temperature of the cryostat is set to -21 °C. (B) The tibialis anterior muscle in the freezing solution is mounted on the sample holder. (C) Sections of 14 µm thickness are attached to glass microscope slides. Please click here to view a larger version of this figure.
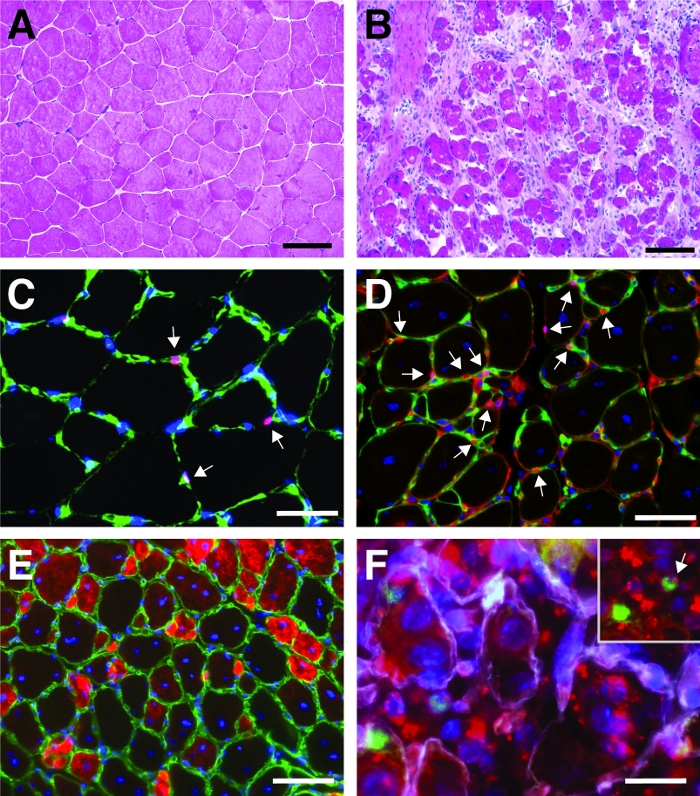
Figure 4: Representative images of regenerating tibialis anterior muscles. H&E staining of tibialis anterior muscles before (A) and 7 days after cardiotoxin injury (B). (C) In uninjured muscle Pax7 positive (in red) satellite cells are located underneath the basal lamina marked by laminin (in green). Nuclei are counterstained with DAPI (in blue). Pax7 positive cells are marked by an arrow. (D) After 10 days of regeneration, myofibers are characterized by centrally located nuclei (in blue), Pax7 is shown in red, laminin in green. Pax7 positive cells are marked by an arrow. (E) Newly formed myofibers express developmental myosin (in red), laminin in green, nuclei are counterstained with DAPI (in blue). (F) Fluorescently labeled self-delivering siRNA (siRED, depicted in red) is still found in satellite cells (Pax7 positive, in green, marked by an arrow in the inset) 2 days after injection of the self-delivering siRNA into regenerating tibialis anterior muscle (injection at day 3 after cardiotoxin mediated injury). Laminin is shown in white, nuclei are counterstained with DAPI (in blue). Scale bar = 100 µm (A and B), 50 µm (C-E), 25 µm (F). Please click here to view a larger version of this figure.
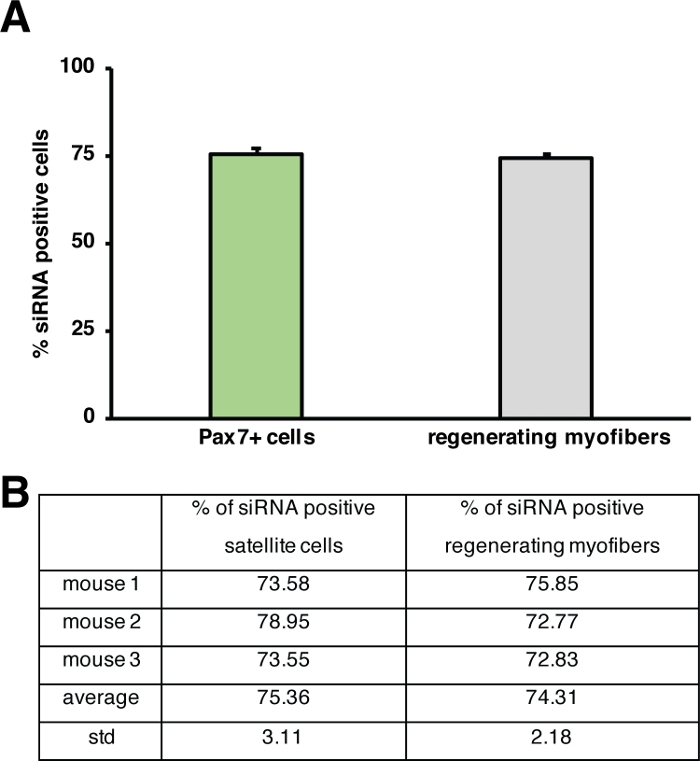
Figure 5: Quantification of uptake of fluorescently labeled siRNA. (A) Quantification of siRNA uptake by satellite cells (Pax7+ cells) and regenerating myofibers 2 days after injection into regenerating skeletal muscle. Injection was performed at day 3 after cardiotoxin mediated injury. n = 3, error bars show SEM. (B) Quantification underlying the graph depicted in (A). Please click here to view a larger version of this figure.
