Sample Preparation using a Lipid Monolayer Method for Electron Crystallographic Studies
Summary
Lipid monolayers have been used as a foundation for forming two-dimensional (2D) protein crystals for structural studies for decades. They are stable at the air-water interface and can serve as a thin supporting material for electron imaging. Here we present the proven steps on preparing lipid monolayers for biological studies.
Abstract
Electron crystallography is a powerful tool for high-resolution structure determination. Macromolecules such as soluble or membrane proteins can be grown into highly ordered two-dimensional (2D) crystals under favorable conditions. The quality of the grown 2D crystals is crucial to the resolution of the final reconstruction via 2D image processing. Over the years, lipid monolayers have been used as a supporting layer to foster the 2D crystallization of peripheral membrane proteins as well as soluble proteins. This method can also be applied to 2D crystallization of integral membrane proteins but requires more extensive empirical investigation to determine detergent and dialysis conditions to promote partitioning to the monolayer. A lipid monolayer forms at the air-water interface such that the polar lipid head groups remain hydrated in the aqueous phase and the non-polar, acyl chains, tails partition into the air, breaking the surface tension and flattening the water surface. The charged nature or distinctive chemical moieties of the head groups provide affinity for proteins in solution, promoting binding for 2D array formation. A newly formed monolayer with the 2D array can be readily transfer into an electron microscope (EM) on a carbon-coated copper grid used to lift and support the crystalline array. In this work, we describe a lipid monolayer methodology for cryogenic electron microscopic (cryo-EM) imaging.
Introduction
Electron diffraction through 2D crystals or helical arrays of proteins can achieve sub-nanometer resolutions in favorable cases1,2,3. Of particular interest are reconstituted 2D membrane protein arrays or crystals in their near-native environments1. Because a crystal acts as a signal amplifier enhancing the intensities of the structural factors at specific spatial frequencies, electron crystallography allows probing a target with a smaller size at high resolutions, such as small molecules, than those for single-particle cryo-EM. The electron beam can be diffracted by an ordered 2D array of proteins, generating a diffraction pattern or a lattice image depending on where the image plane is recorded on the detector4. The diffracted intensities can then be extracted and processed to reconstruct a 2D projection structure of the crystal. Electrons have a larger scattering cross-section than X-rays and its scattering mostly follows the Rutherford model based on the Coulomb interaction between the electrons and the charged atoms in the molecule5. The thicknesses of 2D membrane crystals are usually less than 100 nm, suitable for electron transmission without dynamical scattering occurring within specimen6. Electron crystallographic studies have been shown to be a powerful tool to probe high-resolution structural information of membrane proteins and lipid-protein interactions7,8,9,10,11,12,13,14,15,16,17.
A lipid monolayer is one single lipid layer composed of phospholipids densely packed at an air-water interface6, which can assist the 2D array formation for soluble proteins or peripheral membrane proteins18. Depending on the density of the lipids and their lateral pressure, the lipid molecules can form an ordered 2D array on the air-water interface with their acyl chains extended to the air and hydrophilic headgroups exposed in the aqueous solution1,6,19. The lipid headgroup can interact with proteins via electrostatic interaction or can be modified to provide an affinity tag to bind a specific protein domain. For example, the DOGS-NTA-Ni (1,2-dioleoyl-sn-glycero-3-[(N-(5-amino-1-carboxypentyl)iminodiacetic acid)succinyl]2- Ni2+) is often used in forming a lipid monolayer to bind the proteins with a poly-histidine tag20,21,22. Also, the cholera toxin B can bind a particular pentasaccharide of ganglioside GM1 in a lipid monolayer for structural studies23,24. By anchoring the proteins on the lipid headgroups, the lipid monolayer can assist the formation of the 2D arrays that are thin for high-resolution electron crystallographic studies. The lipid monolayer technique has been used in electron crystallography for structural studies of proteins, such as streptavidin2,25, annexin V26, cholera toxin27, E. coli gyrase B subunit28, E. coli RNA polymerase25,29,30, carboxysome shell proteins31 and the capsid proteins of the HIV-132 and Moloney murine leukemia virus33. Due to the stability and chemical property of the lipid monolayer, different applications for sample preparation have been explored for cryo-EM imaging34. However, optimization will be needed for protein array formation.
Here, we provide extensive details of the general preparation of lipid monolayers for cryo-EM imaging and some considerations that could affect the quality of the formed monolayers.
Protocol
1. Teflon block preparation
- Prepare the Teflon block from chemical-resistant PTFE (polytetrafluoroethylene) resin. Make holes on the block using a general drill followed by the dimensions labeled in Figure 1.
2. Monolayer lipid preparation
NOTE: Estimated operating time: 30- 45 minutes
- Lipid stock preparation
- Prepare a 0.01 mg/mL lipid mixture in 9:1 (v/v) chloroform/methanol using 8.91 mL of chloroform, 0.99 mL of methanol and 0.01 mL of 10 mg/mL lipids.
- Teflon plate preparation
- Sonicate a Teflon plate in a methanol bath for 5 minutes.
- Wash with hot water for 15 minutes.
- Rinse with distilled water.
- Dry the Teflon plate in a desiccator for 30 minutes and keep it under vacuum when not in use (Figure 1).
3. Formation of lipid monolayer on buffer reservoir
NOTE: Estimated operating time: 1.5 hours
- Freshly prepare a lipid mixture of 0.01 mg/mL in a 9:1 mixture of chloroform/methanol on the day of the experiment.
- Place a Type-1 filter paper in a Petri dish and arrange the Teflon block on top of the filter paper.
- Fill the wells of the Teflon plate with 60 µL of the buffer.
- Carefully add lipid mixture on top of the buffer surface drop by drop (ideally 1 µL per drop) using a Hamilton syringe.
- Wet the filter paper with distilled water to keep humidity in the Petri dish.
- Incubate at room temperature for 60 minutes. The chloroform should evaporate, and a monolayer of lipid on the surface of the buffer should be formed.
4. Application of an EM grid on a lipid monolayer
NOTE: Estimated operating time: 1.5 hours
- Gently place a holey carbon-coated EM grid without glow-discharging, carbon side down, onto the top of each buffer reservoir.
- Carefully inject proteins into the side injection well. Use final protein concentrations in the well around 1 µM of protein as a starting point. Pre-optimize the protein concentration before the experiment.
- Incubate for 60 minutes at room temperature.
- Gently inject approximately 20-30 µL of buffer into the side injection port. This will raise the grid above the surface of the Teflon block.
- Immediately pick up the grid with a tweezer and lift it vertically off the droplet.
- Prepare monolayer samples for negative staining or cryo-TEM imaging (Figure 2). If the proteins form a 2D array, the image power spectrum or Fourier periodogram will show diffraction intensities at the spatial frequencies based on the crystal symmetry. If no diffraction spots are shown in the power spectrum, the proteins are unlikely to form a 2D crystal.
- Check the coverage of the lipid monolayer on the EM grid using a transmission electron microscope (TEM). A lower-magnified electron image is helpful to identify the morphology of the covered monolayer. Usually, the image contrast of the monolayer-covered area will be slightly lower than that of the uncovered areas. In addition, the computed 2D image power spectrum will help identify the location of the 2D crystalline.
Representative Results
A lipid monolayer deposited on the EM grid can be visualized under a transmission electron microscope (TEM) without staining. The monolayer presence can be recognized by the contrast difference from the area without any specimen in the beam path. Areas that have lipid monolayer coverage have lower local contrast than the ones with no coverage, since the electron beam through the empty holes has no scattering and shows a brighter illumination (Figure 3).
To screen the conditions for 2D monolayer crystallization, investigate the image of the specimen using negative-stain EM or cryo-EM imaging. Whether the monolayer covers the holes could be observed by a stronger edge contrast when the stage is tilted. If a crystal is formed, the image power spectrum or periodogram will show the Fourier or diffraction spots. Crystals in small sizes (< 200 nm) and randomly oriented may show high-intensity rings in the power spectrum. If the crystals are not grown, the power spectrum will not show Fourier spots.
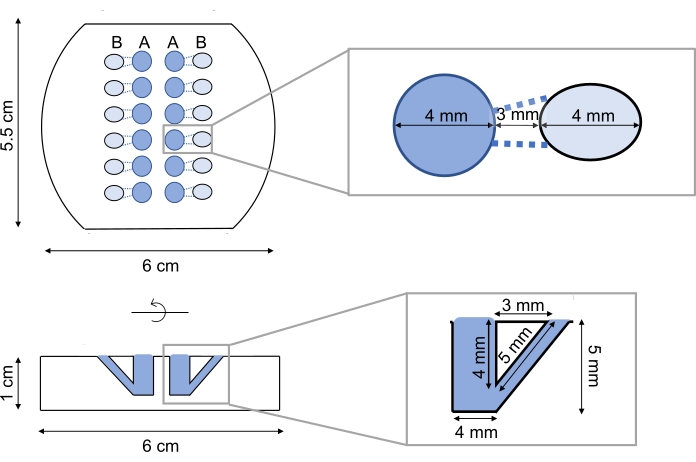
Figure 1. Teflon block used for lipid monolayer formation. A Teflon block is shown in top (above) and cross-section (below) views. It contains 12 wells arranged in six rows and two columns. The main buffer well labeled A, shown in darker blue, in the middle is 4 mm in diameter and 5 mm deep with 60 µL sample volumes. The monolayer is formed on top of the buffer reservoir in the main well and proteins are injected through the side port labeled B shown in light blue. Please click here to view a larger version of this figure.
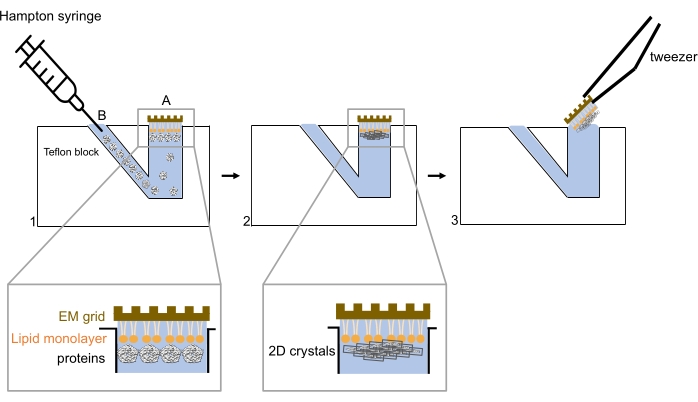
Figure 2. Schematic presentation of the development of 2D crystal on lipid monolayer. The buffer reservoir is first filled with the buffer, followed by the lipid deposited on top of the well. The carbon side of an EM grid is placed on the lipid monolayer, which will interact with the hydrophobic tails. First, proteins are injected through the side injection well B (step 1). Over time, the proteins are arranged into a 2D crystal lattice and interact with the lipid head groups (step 2). To pick up the EM grid for further analysis, a tweezer is used to lift the grid off the buffer reservoir vertically (step 3). Please click here to view a larger version of this figure.
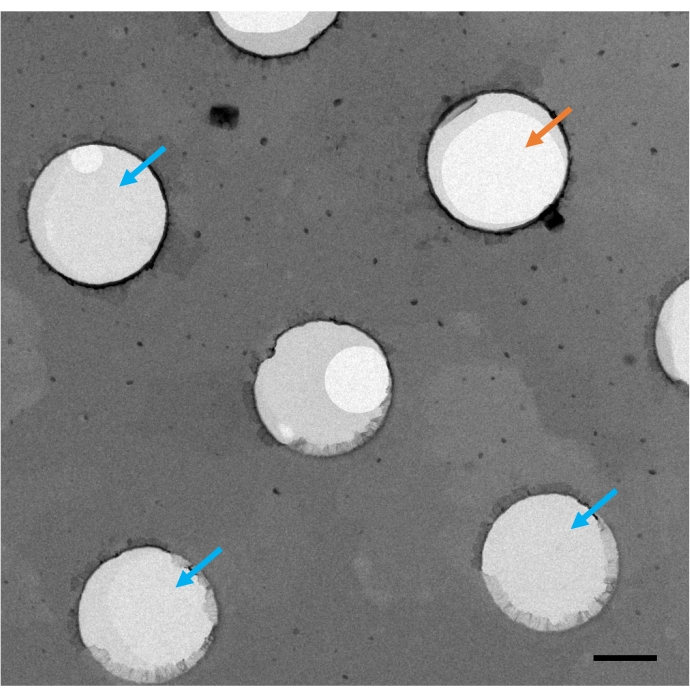
Figure 3. Example of the lipid monolayer coverage on an EM grid. A monolayer- and holey carbon-coated EM grid without staining was imaged using a Philips CM12 transmission electron microscope at room temperature. The orange arrow indicates the area that has no lipid monolayer coverage, and the blue arrows indicate the full coverage of the lipid monolayer in the grid hole. Scale bar is 0.5 μm. Please click here to view a larger version of this figure.
Discussion
A lipid monolayer is a powerful tool that facilitates the growth of large 2D crystals for structural studies of biological macromolecules. To successfully prepare an intact lipid monolayer at the air-water interface, it is strongly recommended that the lipids are prepared freshly on the day of the experiment, because oxidization of the lipid acyl chain could lead to packing disruption in the monolayer and adversely affect the resulting crystal formation. Purchased lipids in powder form should be dissolved using a mixture of chloroform:methanol solvent, which is commonly used to dissolve phospholipids35. The chloroform:methanol solvent can be premade and stored in a sealed glass vial. While the dissolved lipids are applied at the air-water interface, wait until the chloroform:methanol solvent completely evaporates from the buffer. Proper waiting time for the formation of lipid monolayer and the formation of 2D crystals is essential in this experiment. Typically, the formation of a lipid monolayer and 2D arrays can take place within approximately 30 to 60 minutes of incubation time18.
When a lipid monolayer is properly formed, ensure that the 2D crystal array formation can be promoted by the lipid effectively. The buffer composition should be optimized to stabilize the establishment of 2D arrays, especially when DOGS-NTA-Ni lipid is used18,36. The success of the lipid monolayer crystallization for this lipid may be affected if there are high concentrations of EDTA (ethylenediaminetetraacetate), DTT (1,4-dithiothreitol), or various ions in the buffer. The soluble chelator EDTA can disrupt the lipid-metal ion-protein complex36. One example of this is the streptavidin protein interaction with iminodiacetate-Cu (II) lipid; the addition of EDTA removes Cu (II) and interrupts the lipid-metal ion-protein complex20. Hence, the sample may need to be diluted or buffer exchanged to remove EDTA, allowing a stronger binding of protein and promoting crystal formation. The optimization of the buffer needs to be done before it is added to the buffer reservoir in the Teflon block.
To preserve the high-resolution structural detail of the 2D crystal, maintaining the specimen flatness is critical. However, due to surface tension, differential heat capacities between the specimen and support or human intervention, it is not trivial to maintain the flatness of the 2D crystal during the sample transfer. When the EM grid is lifted from the buffer reservoir, one must operate it with as little disturbance as possible to leave the monolayer interface intact. Any drastic rotation or movement of the grid may fracture the monolayer interface. This also applies for sample freezing steps, where the grid with crystals must be handled with care and rapidly frozen to avoid ice crystals, which can damage the newly formed 2D protein crystals.
One may use a lacey carbon-coated grid to support monolayers in different sizes, depending on the hole sizes on the lacey carbon film. The flatness of the monolayer interface may be affected by various hole sizes on the lacey carbon film, which may help screen a flat monolayer with the diffraction intensities in isotropic directions.
There are four common methods used to grow and develop 2D crystals for structural studies: dialysis, dilution, hydrophobic adsorption and lipid monolayer37,38,39. The dialysis method removes the detergents that are used to solubilize membrane proteins in the early step of purification. The 2D crystals are grown by reconstituting detergent-solubilized membrane proteins into lipid bilayers under favorable conditions35. For the reconstitution to take place, detergents need to be removed by dialysis. Therefore, the dialysis method helps to drive the formation of 2D crystals40. However, the dialysis method is usually time-consuming, especially for detergents with a small critical micelle concentration (CMC). The formation of 2D arrays can also be achieved via a controlled dilution. This method provides a set ratio for the protein-lipid-detergent mixture to induce crystal formation if the detergent concentration drops below its CMC. This could be facilitated by a specific dilution machine to carry out the work starting with a higher initial protein concentration41. In addition, hydrophobic adsorption prompts the growth of 2D crystals by using polystyrene beads (e.g., Bio-beads) to remove the low CMC detergents42. If the protein of interest is solubilized with high CMC detergents, this method is not ideal. In contrast, the lipid monolayer method is versatile and provides non-convoluted steps for 2D crystallization43. As discussed here, it requires a simple apparatus and some materials to conduct the experiments. With our method, the total time needed for the experiment can be shortened to 4 hours17. Even though pre-optimization of certain parameters, such as protein concentration and buffer components, may still be required beforehand, it is possible to maximize one's efficiency in routine lab work.
For crystallizing integral membrane proteins using a lipid monolayer, one should test whether the detergent concentration for solubilizing protein and lipids disturbs the monolayer interface. Generally, it is suggested to use the detergent concentration lower than the CMC 44. In cases that the monolayer interface is sensitive to the detergent existence, an alternative approach can be to form the lipid monolayer using fluoro-lipids, which are hardly solubilized by detergent and expand the applicability of integral membrane protein crystallization using the monolayer method45.
Although the lipid monolayer method is simple and less costly, the reproducibility of the monolayer preparation depends on various factors, such as ambient humidity and temperature. The success of the sample transfer for cryo-EM imaging relies on careful specimen handling. In some cases, it may be challenging to obtain high-resolution results by a novice or in the beginning phase of a new research project.
In conclusion, the lipid monolayer method offers an opportunity for studying protein structures due to its simplicity. It may provide an alternative approach to facilitate the process of 2D crystal formation.
Disclosures
The authors have nothing to disclose.
Acknowledgements
The preparation of this manuscript was partially supported by US Army Research Office (W911NF2010321) and Arizona State University startup funds to P.-L.C.
Materials
| 14:0 PC (DMPC) | Avanti Lipids | 850345 | 1,2-dimyristoyl-sn-glycero-3-phosphocholine, 1 x 25 mg, 10 mg/mL, 2.5 mL |
| Bulb for small pipets | Fisher Scientific | 03-448-21 | |
| Chloroform | Sigma-Aldrich | C2432 | |
| Desiccator vacuum | Southern Labware | 55207 | |
| EM grids | Electron Microscopy Sciences | CF413-50 | CF-1.2/1.3-4C 1.2 µm hole, 1.3 µm space |
| Filter paper | GE Healthcare Life Sciences | 1001-090 | Diameter 90 mm |
| Glass Pasteur pipets | Fisher Scientific | 13-678-20A | |
| Hamilton syringe (25 µL) | Hamilton Company | 80465 | |
| Hamilton syringe (250 µL) | Hamilton Company | 81165 | |
| Hamilton syringe (5 µL) | Hamilton Company | 87930 | |
| Hamilton syringe (500 µL) | Hamilton Company | 203080 | |
| Methanol | Sigma-Aldrich | M1775-1GA | |
| Petri dish | VWR | 25384-342 | 100 mm × 15 mm |
| Teflon block | Grainger | 55UK05 | 60 µL wells with side injection ports, manually made |
| Tweezers | Electron Microscopy Sciences | 78325 | Various styles |
| Ultra-pure water | |||
| Ultrasonic cleaner | VWR | 97043-996 |
References
- Raunser, S., Walz, T. Electron crystallography as a technique to study the structure on membrane proteins in a lipidic environment. Annual Review of Biophysics. 38 (1), 89-105 (2009).
- Avila-Sakar, A. J., Chiu, W. Visualization of beta-sheets and side-chain clusters in two-dimensional periodic arrays of streptavidin on phospholipid monolayers by electron crystallography. Biophysical Journal. 70 (1), 57-68 (1996).
- Braun, T., Engel, A. Two-dimensional electron crystallography. Nature Encyclopedia of Life Sciences. , (2004).
- Wang, H. -. W., Wang, J. -. W. How cryo-electron microscopy and X-ray crystallography complement each other. Protein Science: a publication of the Protein Society. 26 (1), 32-39 (2017).
- Williams, D. B., Carter, C. B. . Transmission electron microscopy. , (2016).
- Abeyrathne, P. D., et al. 1.15 Analysis of 2-D Crystals of Membrane Proteins by Electron Microscopy. Comprehensive Biophysics. , 277-310 (2012).
- Muller, M. P., et al. Characterization of Lipid-Protein Interactions and Lipid-Mediated Modulation of Membrane Protein Function through Molecular Simulation. Chemical Reviews. 119 (9), 6086-6161 (2019).
- Martínez-Ballesta, M. D. C., Carvajal, M. Mutual Interactions between Aquaporins and Membrane Components. Frontiers in Plant Science. 7, 1322 (2016).
- Hite, R. K., Chiu, P. -. L., Schuller, J. M., Walz, T. Effect of lipid head groups on double-layered two-dimensional crystals formed by aquaporin-0. PloS One. 10 (1), 0117371 (2015).
- Murata, K., et al. Structural determinants of water permeation through aquaporin-1. Nature. 407 (6804), 599-605 (2000).
- Schenk, A. D., et al. The 4.5 A structure of human AQP2. Journal of Molecular Biology. 350 (2), 278-289 (2005).
- Gonen, T., et al. Lipid-protein interactions in double-layered two-dimensional AQP0 crystals. Nature. 438 (7068), 633-638 (2005).
- Hiroaki, Y., et al. Implications of the aquaporin-4 structure on array formation and cell adhesion. Journal of Molecular Biology. 355 (4), 628-639 (2006).
- Gonen, T., Sliz, P., Kistler, J., Cheng, Y., Walz, T. Aquaporin-0 membrane junctions reveal the structure of a closed water pore. Nature. 429 (6988), 193-197 (2004).
- Chiu, P. -. L., et al. The structure of the prokaryotic cyclic nucleotide-modulated potassium channel MloK1 at 16 A resolution. Structure. 15 (9), 1053-1064 (2007).
- Kowal, J., et al. Ligand-induced structural changes in the cyclic nucleotide-modulated potassium channel MloK1. Nature Communications. 5, 3106 (2014).
- Walz, T., Grigorieff, N. Electron Crystallography of Two-Dimensional Crystals of Membrane Proteins. Journal of Structural Biology. 121 (2), 142-161 (1998).
- Yeager, M., Dryden, K. A., Ganser-Pornillos, B. K. Lipid monolayer and sparse matrix screening for growing two-dimensional crystals for electron crystallography: methods and examples. Methods in Molecular Biology. 955, 527-537 (2013).
- Pal, S. Chapter 6 – Structure analysis and visualization. Fundamentals of Molecular Structural Biology. , 119-147 (2020).
- Frey, W., et al. Two-dimensional protein crystallization via metal-ion coordination by naturally occurring surface histidines. Proceedings of the National Academy of Sciences of the United States of America. 93 (10), 4937-4941 (1996).
- Kubalek, E. W., Le Grice, S. F., Brown, P. O. Two-dimensional crystallization of histidine-tagged, HIV-1 reverse transcriptase promoted by a novel nickel-chelating lipid. Journal of Structural Biology. 113 (2), 117-123 (1994).
- Vénien-Bryan, C., et al. Structural study of the response regulator HupR from Rhodobacter capsulatus. Electron microscopy of two-dimensional crystals on a nickel-chelating lipid. Journal of Molecular Biology. 274 (5), 687-692 (1997).
- Merritt, E. A., Sarfaty, S., vanden Akker, F., L’Hoir, C., Martial, J. A., Hol, W. G. Crystal structure of cholera toxin B-pentamer bound to receptor GM1 pentasaccharide. Protein Science: a publication of the Protein Society. 3 (2), 166-175 (1994).
- Mosser, G., Mallouh, V., Brisson, A. A 9 A two-dimensional projected structure of cholera toxin B-subunit-GM1 complexes determined by electron crystallography. Journal of Molecular Biology. 226 (1), 23-28 (1992).
- Edwards, A. M., Darst, S. A., Hemming, S. A., Li, Y., Kornberg, R. D. Epitaxial growth of protein crystals on lipid layers. Nature Structural Biology. 1 (3), 195-197 (1994).
- Olofsson, A., Mallouh, V., Brisson, A. Two-dimensional structure of membrane-bound annexin V at 8 A resolution. Journal of Structural Biology. 113 (3), 199-205 (1994).
- Ribi, H. O., Ludwig, D. S., Mercer, K. L., Schoolnik, G. K., Kornberg, R. D. Three-dimensional structure of cholera toxin penetrating a lipid membrane. Science. 239 (4845), 1272-1276 (1988).
- Celia, H., et al. Three-dimensional model of Escherichia coli gyrase B subunit crystallized in two-dimensions on novobiocin-linked phospholipid films. Journal of Molecular Biology. 236 (2), 618-628 (1994).
- Darst, S. A., Kubalek, E. W., Kornberg, R. D. Three-dimensional structure of Escherichia coli RNA polymerase holoenzyme determined by electron crystallography. Nature. 340 (6236), 730-732 (1989).
- Schultz, P., et al. Structural study of the yeast RNA polymerase A. Electron microscopy of lipid-bound molecules and two-dimensional crystals. Journal of Molecular Biology. 216 (2), 353-362 (1990).
- Dryden, K. A., Crowley, C. S., Tanaka, S., Yeates, T. O., Yeager, M. Two-dimensional crystals of carboxysome shell proteins recapitulate the hexagonal packing of three-dimensional crystals. Protein Science: a publication of the Protein Society. 18 (12), 2629-2635 (2009).
- Barklis, E., McDermott, J., Wilkens, S., Fuller, S., Thompson, D. Organization of HIV-1 capsid proteins on a lipid monolayer. The Journal of BIOLOGICAL CHemistry. 273 (13), 7177-7180 (1998).
- Barklis, E., et al. Structural analysis of membrane-bound retrovirus capsid proteins. The EMBO Journal. 16 (6), 1199-1213 (1997).
- Kelly, D. F., Dukovski, D., Walz, T. Monolayer purification: a rapid method for isolating protein complexes for single-particle electron microscopy. Proceedings of the National Academy of Sciences of the United States of America. 105 (12), 4703-4708 (2008).
- Reis, A., Rudnitskaya, A., Blackburn, G. J., Mohd Fauzi, N., Pitt, A. R., Spickett, C. M. A comparison of five lipid extraction solvent systems for lipidomic studies of human LDL. Journal of Lipid Research. 54 (7), 1812-1824 (2013).
- Ueda, E. K. M., Gout, P. W., Morganti, L. Current and prospective applications of metal ion-protein binding. Journal of Chromatography. A. 988 (1), 1-23 (2003).
- Dietrich, J., nien-Bryan, C. . Strategies for Two-dimensional Crystallization of Proteins Using Lipid Monolayers. , (2005).
- Kuang, Q., Purhonen, P., Hebert, H. Two-Dimensional Crystallization Procedure, from Protein Expression to Sample Preparation. BioMed Research International. 2015, 693869 (2015).
- De Zorzi, R., Nicholson, W. V., Guigner, J. -. M., Erne-Brand, F., Vénien-Bryan, C. Growth of large and highly ordered 2D crystals of a K+ channel, structural role of lipidic environment. Biophysical Journal. 105 (2), 398-408 (2013).
- Johnson, M. C., Schmidt-Krey, I. Two-dimensional crystallization by dialysis for structural studies of membrane proteins by the cryo-EM method electron crystallography. Methods in Cell Biology. 113, 325-337 (2013).
- Rémigy, H. -. W., Caujolle-Bert, D., Suda, K., Schenk, A., Chami, M., Engel, A. Membrane protein reconstitution and crystallization by controlled dilution. FEBS Letters. 555 (1), 160-169 (2003).
- Braun, T., Kaufmann, T. C., Rémigy, H., Engel, A. Two-dimensional Crystallization of Membrane Proteins. Encyclopedic Reference of Genomics and Proteomics in Molecular Medicine. , 1936-1942 (2006).
- Lebeau, L., Vénien-Bryan, C. Monolayer two-dimensional crystallization of membrane proteins. Methods in Molecular Biology. 955, 59-71 (2013).
- Seddon, A. M., Curnow, P., Booth, P. J. Membrane proteins, lipids and detergents: not just a soap opera. Biochimica et Biophysica Acta. 1666 (1-2), 105-117 (2004).
- Lebeau, L., et al. Two-dimensional crystallization of a membrane protein on a detergent-resistant lipid monolayer. Journal of Molecular Biology. 308 (4), 639-647 (2001).
