Kesin gen düzenleme genetik hastalıklar mücadele için yeni strateji geliştirme dahil önemli Biyomedikal gelişmeler dönüm noktası oluşturur. At gen düzenleme teknolojileri ön planda olduğunu lustered c regularly –ınterspaced short palindromic repeats (CRISPR) kullanımı hakkında güvenerek yöntemi / başlangıçta tespit edildi Cas9 sistemi Bakteriyel bağışıklık (başvurular1,2‘ gözden) viral genetik materyal işgali karşı bir bileşeni olarak. Çinko-parmak enzimler (ZFNs) ve (başvuru3gözden geçirme), transkripsiyon harekete geçirmek gibi efektör enzimler (TALENs) gibi diğer gen düzenleme araçları üzerinde büyük bir avantaj CRISPR/Cas9 sisteminin göreli basitliğidir plazmid tasarım ve İnşaat CRISPR bileşenlerinin — gen düzenleme birkaç özel laboratuvarları üzerinden daha geniş bir araştırma topluluk sadece bir genişleme powered bir özellik. Ayrıca, CRISPR/Cas9 programlama sadeliği ve çoğaltılmış hedef tanıma kapasitesini daha da popülaritesini düşük maliyetli ve kolay kullanımlı teknoloji olarak yol açtı. Araştırmacılar bu tür gen düzenleme bileşenleri hücrelere sunmak için kullanabileceği çeşitli yöntemler arasında viral vektörler farkla en popüler ve etkili sistem kalır.
Lentiviral vektörel çizimler (LVs) CRISPR/Cas9 sistem vivo içinde çeşitli uygulamalar4,5,6,7için bileşenleri sunmak için Seçim aracı olarak ortaya çıkmıştır. Birkaç temel özellikleri LVs hücrelerin bölünmesi ve sigara bölen, düşük immünojenisite hem en az hücresel toksisite (başvuru8‘ gözden) bulaştırmak onların da dahil olmak üzere bu işlem için popüler bir seçimdir. Sonuç olarak, LV-aracılı gen tedavisi tedaviler HIV-1, HBV ve HSV-1, gibi bulaşıcı hastalıkların yanı sıra insan kalıtsal hastalıklar, Kistik fibrozis ve neo-vasküler makula dejenerasyonu gibi temel hataları düzeltme istihdam edilmiştir 4 , 5 , 7 , 9 , 10 , 11. Ayrıca, LVs etkili bir şekilde değiştirilmiş bir tek vektör sistemi12kullanarak farklı genomik loci multiplex gen düzenleme işlemleri yapmak için.
Ancak, ana bilgisayar genom entegre LVs doğal özellik ve mutajenik olabilir sık sık onların programı transgene teslim araçlarda, özellikle klinik ayarları olarak yetersizlik. LVs stabil entegre sürdürülebilir yüksek düzeylerde onların transgenes hızlı, Ayrıca, bu sistem kötü CRISPR/Cas9 gibi gen düzenleme bileşenlerinin teslimatlar için uygun olduğu için; overexpression Cas9-guide RNA (gRNA) ve ZFNs gibi benzer proteinler yüksek seviyede istenmeyen mutasyonlar13,14,15,16 dahil hedef kapalı etkileri ile ilişkili , 17 ve potansiyel olarak sitotoksisite18geliştirebilirsiniz. Bu nedenle, kesin ulaşmak için gen düzenleme ile en az hedef sonuç, bu geçici Ifade gen bileşenlerinin düzenlenmesine izin tasarım sistemleri için zorunludur.
Son yıllarda, farklı platformlarda teslim geliştirilmiştir geçici hücreleri16,19,20,21 (başvuru22gözden) CRISPR/Cas9 ifade etmek için. Bunlar doğrudan uygun Kılavuzu RNA’lar ile birlikte saf Cas9 hücrelere, hangi hedeflenen gen düzenleme plazmid aracılıklı transfection16ile karşılaştırıldığında, daha etkili olduğu gösterilmiştir tanıtımı üzerinde itimat yöntemleri içerir. Çalışmalar bu Ribonükleoprotein (RNP) göstermiştir oluşan komplekslerin rehberlik RNA/Cas9 parçacıklar hızla devre DNA bölünme bu bileşenlerin kısa vadeli ifade ulaşmak için yeterli olduğunu düşündüren hedeflerine, arabuluculuk sonra sağlam gen16düzenleme. Makul, entegre adeno ilişkili viral vektörel çizimler (AAVs) gibi viral vektör platformlar hücrelere gen düzenleme makine sunmak için bir alternatif sağlayabilir. Ne yazık ki, önemli ölçüde daha düşük ambalaj yeteneği LVs daha AAV capsids sahip (< 5kb), hangi ciddi çok bileşenli CRISPR toolkit (başvuru8‘ gözden) tek bir vektör içinde paketlemek için yeteneklerini engellemektedir. Histon uyku (örneğin, sodyum bütrat23) inhibe veya hücre döngüsü (örneğin, kafein24) engel bileşikler ilavesi lentiviral titreleri artırmak için gösterilmiş olduğunu fazlalaştı. Son rağmen defa geliştirilen geçici ifade sistemleri hala azaltılmış viral titreleri ve düşük iletim verimliliği ile oluşturulan virüslerin yol birkaç eksiklikler, daha düşük üretim verimliliği gibi tarafından engellemiştir Böyle25yaklaşıyor.
AAV benzeri episomal bakım hücrelerdeki yararı ile LVs ambalaj kapasitesini AEI Integrase-eksik lentiviral vektörler (IDLVs) gen teslim araçların geliştirilmesinde önemli bir ilerleme temsil eder. Bu özellikler IDLVs büyük ölçüde vektörel çizimler, vis-à-vis sürekli overexpression olası genotoksik öğeleri ve entegrasyon-aracılı mutagenisitesinin entegre ile ilişkili önemli sorunları aşmak yardımcı olur. Daha önce IDLVs başarıyla episomal gen ifade26,27geliştirmek için değiştirilebilir gösterilmiştir. IDLV-aracılı CRISPR/Cas9 teslim açısından düşük üretim titreleri ve episome kaynaklı genleri integrase usta lentiviral sistemleri göre alt ifade sınırlar genom düzenleme teslim etmek için bona fide araçları olarak onların yardımcı programı transgenik yapıları. Biz son zamanlarda transgene ifade ve viral titreleri IDLV üretimle ilişkili önemli ölçüde siteleri viral ifade kaset28içinde transkripsiyon faktörü Sp1 için bağlama dahil tarafından geliştirilmiş olduğunu gösterdi. Değiştirilmiş IDLVs sağlam CRISPR-aracılı gen (HEK-293T hücrelerdeki) vitro ve in vivo (içinde sonrası Mitotik beyin sinir hücreleri), ICLV-aracılı ilgili karşılaştırıldığında çok az hedef kapalı mutasyonlar inducing düzenleme desteklenen sistemleri28. Genel olarak, bir roman geliştirdiğimiz, kompakt, tüm-içinde-bir CRISPR toolkit bir IDLV platformu üzerinde taşınan ve gelişmiş gen düzenleme gibi teslim araç kullanmanın çeşitli yararları sıraladı.
Burada, IDLV-CRISPR/Cas9 sisteminin üretim protokolü, derleme, arıtma, konsantrasyon ve titrasyon IDLVs, hem de stratejileri bu vektörel çizimler gen düzenleme etkinliğini doğrulamak için çeşitli adımlar da dahil olmak üzere tanımlanır. Bu iletişim kuralı farklı müfettişler ihtiyaçlarını karşılamak için kolayca ölçeklenebilir ve başarıyla birimleri (TU) transducing 1 x 1010 aralıkta titreleri ile LV vektörel çizimler oluşturmak için tasarlanmıştır / mL. Bu protokolü ile oluşturulan vektörel çizimler verimli bir şekilde birkaç farklı hücre tipleri, zor transduce embriyonik kök hücreler, hematopoetik hücreler (T-hücreleri ve makrofajlar) dahil olmak üzere ve kültürlü ve içinde vivobulaştırmak için kullanılması gereken- enjekte nöronlar. Ayrıca, protokol eşit integrase yetkili lentiviral vektörler benzer miktarlarda üretimi için uygundur.
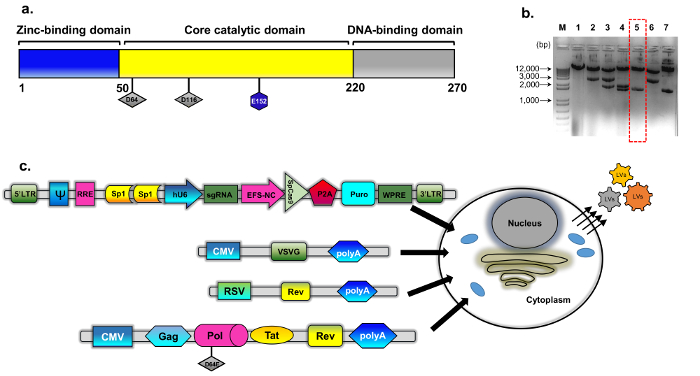
Şekil 1: IDLV ambalaj. (a) şeması değiştirilmiş plazmid psPAX2 elde edilen vahşi türü integrase protein (b) (bkz: yöntemleri, plazmid inşaat detayları için). Mutasyona uğramış integrase klonlar için ekranlı klonlar temsilcisi özel jel görüntüsü. Bir standart plazmid DNA izolasyon Mini kit kullanılarak hazırlanan DNA örnekleri sindirim EcoRV ve SphI tarafından analiz edildi. Doğru sindirmek klon (sayı 5, kesik çizgili kırmızı kutu) daha fazla doğrudan (Sanger) sıralama INTD64E değişimi için tarafından doğrulandı. İntegrase-eksik ambalaj kaset pBK43 seçildi. (c) 293T hücre transfected VSV-G, ambalaj ve transgene kaset (Sp1-CRISPR/Cas9 tüm-içinde-bir plazmid) ile gösterilen IDLV-CRISPR/Cas9 Vektörler, üretmek için şematik geçici transfection Protokolü’nün istihdam. Hücre zarının bud viral parçacıkların (transgene kaset ifade edilir) vektör tam uzunlukta RNA içerir. Tat düzenleyici proteinler içeren IDLV paket yönetim sistemi ikinci nesil kullanıldı ve Rev Rev ifade daha da bir ayrı kaset (RSV-REV-plazmid) desteklenmiştir. ABBREV: soldan sağa-uzun-terminal tekrar ediyorum, VSV-G, vesicular Stomatit virüs G-protein, pCMV-Sitomegalovirüs organizatörü; Rous sarkomu (RSV) virüs organizatörü; RRE-(Rev yanıt öğesi). Diğer düzenleyici elemanlarının ifade kasetin Sp1 bağlayıcı siteleri, Rev Response öğesi (RRE), dahil Dağ sıçanı hepatit virüsü Posttranscriptional düzenleyici eleman (WPRE), bir çekirdek-uzama faktör 1α organizatörü (EFS-NC), vektör ambalaj öğe ψ (PSI), insan Sitomegalovirüs (hCMV) organizatörü ve insan U6 organizatörü. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
