Lentiviral Mediated Gene Silencing in Human Pseudoislet Prepared in Low Attachment Plates
Summary
A protocol to create gene modified human pseudoislets from dispersed human islet cells that are transduced by lentivirus carrying short hairpin RNA (shRNA) is presented. This protocol utilizes readily available enzyme and culture vessels, can be performed easily, and produces genetically modified human pseudoislets suitable for functional and morphological studies.
Abstract
Various genetic tools are available to modulate genes in pancreatic islets of rodents to dissect function of islet genes for diabetes research. However, the data obtained from rodent islets are often not fully reproduced in or applicable to human islets due to well-known differences in islet structure and function between the species. Currently, techniques that are available to manipulate gene expression of human islets are very limited. Introduction of transgene into intact islets by adenovirus, plasmid, and oligonucleotides often suffers from low efficiency and high toxicity. Low efficiency is especially problematic in gene downregulation studies in intact islets, which require high efficiency. It has been known that enzymatically-dispersed islet cells reaggregate in culture forming spheroids termed pseudoislets. Size-controlled reaggregation of human islet cells creates pseudoislets that maintain dynamic first phase insulin secretion after prolonged culture and provide a window to efficiently introduce lentiviral short hairpin RNA (shRNA) with low toxicity. Here, a detailed protocol for the creation of human pseudoislets after lentiviral transduction using two commercially available multiwell plates is described. The protocol can be easily performed and allows for efficient downregulation of genes and assessment of dynamism of insulin secretion using human islet cells. Thus, human pseudoislets with lentiviral mediated gene modulation provide a powerful and versatile model to assess gene function within human islet cells.
Introduction
The loss of functional beta cell mass is the central pathology for both type 1 and type 2 diabetes1. While beta cells are the producers of insulin in pancreatic islets, communication between beta cells and non-beta cells plays a critical role in the regulation of insulin secretion2. In addition, dysregulation of glucagon secretion contributes to hyperglycemia in diabetes3. Thus, there is strong interest to modulate gene expression of cells within pancreatic islets to address the mechanism behind the development of islet dysfunction in diabetes. A variety of approaches including transgenic mice are available to modulate gene expression of mouse islets. However, human and mouse islets show distinct innervation, cell distribution, ratio of beta to alpha cells, and response to secretagogues4. Therefore, direct assessment of gene function in human islets is extremely important for understanding the pathophysiology of human pancreatic islets.
Adenoviral vector is the most widely used viral vector to transduce pancreatic islets in vitro due to the high efficiency of transduction in non-dividing cells. However, adenovirus does not penetrate to the core of islets efficiently, especially in human islets5, and is cytotoxic at high doses6. Comparatively, lentiviral vector is less cytotoxic and delivers exogenous genes permanently into the chromosome of post-mitotic cells, making it a widely tested vehicle for gene therapy7. However, the ability of the lentivirus to penetrate the core of intact human islets is also limited, thus requiring partial dispersion by enzymatic digestion to increase the transduction efficiency8. The caveat with the dispersion of intact human islets is the interruption of cell-cell and cell-matrix communication, which compromises the dynamic regulation of insulin secretion critical for the maintenance of glucose homeostasis in humans9. Thus, it has been challenging to assess the impact of gene modulation on the dynamic regulation of islet function in a model of human islets.
It has been known that dispersed islet cells from human and rodent islets autonomously reaggregate into islet-like structures called “pseudoislets”. Pseudoislets show beta and non-beta cell distribution similar to native islets10,11. Additionally, after long-term culture, native islets progressively lose robust first phase insulin secretion5,10,11,12. Yet, pseudoislets demonstrated better preservation of first phase insulin secretion in response to glucose compared with native islets after the same culture period5. In addition to having better preservation of insulin secretion, size-controlled reaggregation of human islet cells in low attachment plates11 provides a window of opportunity to introduce lentivirus vectors prior to their reaggregation into pseudoislets. Several studies have demonstrated the utility of pseudoislets combined with lentiviral mediated transduction. Caton et al.13 reported that the introduction of the green fluorescent protein (GFP) expressing lentivirus had little effect on insulin secretion while achieving homogenous expression of GFP in rat pseudoislets compared with non-infected control. They also demonstrated the specific effect of different connexins on insulin secretion by overexpressing connexins 32, 36, and 43 via lentivirus13. Human pseudoislets prepared with a commercially available 96-well ultra-low attachment plate demonstrated that lentiviral-mediated overexpression of transcription factor SIX3 improves insulin secretion assessed by static incubation14. Recently, human pseudoislets prepared with a 96-well ultra-low attachment plate were used to downregulate glucokinase via lentiviral short hairpin RNA (shRNA) as a proof of principle to show that glucose-stimulated insulin secretion is reduced, while KCl-stimulated insulin secretion was preserved5. The study also demonstrated that human pseudoislets are similar to native islets in gene expression and secretory profiles, further supporting the utility of human pseudoislets to dissect the regulation of islet function5. Although perifusion was not performed, a bioengineered microwell culture plate that recently became commercially available, was also reported to be compatible for lentiviral transduction and produced human pseudoislets that exhibited excellent insulin secretion in vitro and in vivo after transplantation11. Collectively, human pseudoislet formation combined with lentiviral transduction is a simple and efficient approach to investigate human islet pathophysiology, providing a valuable tool to perform mechanistic studies in human islets.
In the current report, a protocol to form human pseudoislets transduced with lentivirus using two commercially available platforms, a 96-well ultra-low attachment plate and a microwell culture plate is presented. Both achieve efficient modulation of gene expression and create human pseudoislets that are compatible for downstream assessments including static incubation and perifusion.
Protocol
Prior to commencement of studies, a human subjects research determination was made by the University of Iowa Institutional Review Board, who determined that the study did not meet the criteria for human subjects research. Consult the local review board before the initiation of the study to determine if the source of islets and planned study requires prior approval.
NOTE: Typically, 1,200−1,400 islet equivalent (IEQ) of human islets are required for the formation of 192 pseudoislets at the size of 3,000 cells/pseudoislets in a 96-well ultra-low attachment plate or 1,200 pseudoislets at the size of 500 cells/pseudoislets in a microwell culture plate. IEQ of islets required varies between different preparations of human islets as donor factors (age, health, weight), isolation efficiency, and culture conditions affect the yield of the single cell suspension. In this protocol, lentivirus containing shRNA targeting a gene of interest is used. The cytomegalovirus (CMV) and human phosphoglycerate kinase (hPGK) promoter based lentiviral vectors are reported to down-regulate gene efficiently in human pseudoislets5,15. The use of lentivirus requires precaution as biohazard16. Contact the local biosafety committee prior to the initiation of the use of lentivirus.
1. Overnight Culture of Human Islets for Recovery After Shipment
- Prepare Connaught Medical Research Laboratories 1066 (CMRL-1066) medium supplemented with 1% human serum albumin (HSA) by combining 50 mL of CMRL-1066, 0.5 g of HSA, 0.5 mL of penicillin-streptomycin, and 0.5 mL of 100 mg/mL glutamine (1% HSA CMRL) in biological safety cabinet (BSC) and passing through a 0.2 μm filter for sterilization.
- Gently swirl the shipping bottle to keep islets in suspension. Transfer the shipping medium containing islets to a 50 mL conical centrifuge tube. Let the tube sit in BSC for 15 min so that islets settle to the bottom of the tube.
- Remove shipping medium gently without disturbing the islet pellet using a 10 mL pipette. Re-suspend the islet pellet in 1% HSA CMRL to a concentration of 400 IEQ/mL.
- Transfer islets into a non-tissue culture treated dish. If islets are split into multiple dishes, keep islets evenly suspended in medium by gently swirling before splitting. Culture islets at 37 °C in a 5% CO2 incubator overnight.
NOTE: The use of a non-tissue culture treated dish is required to prevent the attachment of islets to the plate.
2. Preparation of Single Cell Suspension from Human Islets
- Prepare the following: CMRL-1066 medium with 10% heat-inactivated fetal bovine serum (HiFBS), penicillin-streptomycin, and glutamine (10% HiFBS CRML) at room temperature (RT), a 40 µm strainer, a 35 mm Petri dish, a 1 mL tuberculin syringe, and a hemocytometer.
- Transfer human islets after overnight culture into a 15 mL conical centrifuge tube. Centrifuge at 190 x g for 5 min in a swinging-bucket rotor. Aspirate medium with a 5 mL pipette without disturbing the islet pellet.
- Wash the pellet by adding 10 mL of phosphate-buffered saline (PBS) into the tube, mix gently, and centrifuge at 190 x g for 5 min. Aspirate PBS without disturbing the islet pellet.
- Re-suspend the islet pellet in 0.5 mL of a pre-warmed proteolytic and collagenolytic enzyme mixture and pipette 5x using a P1000 pipette to mix islets. Incubate at 37 °C for 5 min. Mix by pipetting up and down gently 1−5 times.
NOTE: Aggressive pipetting will increase cell loss. - Check for cloudiness (single cells) and the number of flakes (undigested islets). Add 2−3 min to 37 °C incubation depending on the extent of digestion judged by cloudiness and the number of flakes. Stop digestion when flakes are reduced to ~10% of predigestion and solution is cloudy.
NOTE: Time required for digestion differs depending on the islet size distribution of each human islet preparation. - Place a 40 µm strainer in a 35 mm Petri dish and wet the strainer by adding 1 mL of 10% HiFBS CMRL and pressing with 1 mL syringe plunger. Transfer all the cell suspension on top of the strainer and collect the pass-through in a fresh 15 mL tube.
- Wash the tube used for islet digestion with 0.5 mL of fresh CMRL medium to collect leftover cells and pass the wash through the strainer. Combine the pass-through in a 15 mL tube. Repeat once.
- Next, dissociate undigested islets remaining on the strainer by pressing the strainer placed in a 35 mm dish with 1 mL syringe plunger. Collect pass-through again and wash the strainer with fresh CMRL-1066 to remove all remaining digested islets from the strainer and the dish. A total ~3 mL of single cell suspension will now be in the 15 mL tube.
- Record the total volume of the cell suspension and take 10 µL aliquot of cells to count the cell number on a hemocytometer.
- Centrifuge the cell suspension for 5 min at 200 x g. Remove medium without disturbing the pellet. Proceed to step 3.1.1 if using a 96-well ultra-low attachment plate or step 3.2.1 if using a 24 well microwell culture plate to reaggregate the cells.
3. Pseudoislet Formation and Transduction by Lentivirus
- Protocol using a 96-well ultra-low attachment plate
- Determine the desired number of cells per pseudoislet and the number of pseudoislets to create. Typically, 1,000−3,000 cells are used for each pseudoislet for a 96-well ultra-low attachment plate. For 3,000 cells per pseudoislet, adjust the cell suspension to 1 x 105 cells/mL by resuspending the islet pellet from step 2.10 in 10% HiFBS CMRL so that 30 µL of cell suspension has 3,000 cells. Calculate the total volume of 1 x 105 cells/mL of single cell suspension (mL) required using the following equation:
The total volume of 1 x 105 cells/mL of single cell suspension (mL) = (number of cells per pseudoislet) x (number of pseudoislets being made) / 1 x 105.
NOTE: Adjust the concentration of the cell suspension based on the desired number of cells per pseudoislet so that 30 µL of cell suspension makes one pseudoislet. - Transfer the required volume (30 µL x number of pseudoislets being made) of single cell suspension to a fresh 15 mL tube. Add 250 transduction units (TU)/cell of lentivirus containing shRNA targeting a gene of interest or control.
CAUTION: Lentivirus is classified as biosafety level 2 and can be integrated into DNA of infected cells.
NOTE: Use concentrated lentivirus so that the volume of lentivirus added is minimal. Titer required per cell for efficient gene silencing may differ depending on the lentiviral construct. - Mix cell suspension with virus by pipetting gently 5x with a P1000 pipette. Transfer mixed cells into a 50 mL sterile reagent reservoir if using an 8-channel pipette.
- Dispense 30 µL per well of single cell suspension mixed with lentivirus into each well using an 8-channel pipette or a P200 pipette depending on the number of the wells.
- Centrifuge the 96-well plate in a swinging-bucket plate centrifuge at 270 x g at RT for 7 min. Check whether cells are gathered in the center of each well. If not, centrifuge again as gathering of all cells in the center of the well is critical for pseudoislet formation. Culture at 37 °C in a humidified 5% CO2 incubator overnight.
- Add 100 µL of pre-warmed 10% HiFBS CMRL per well next morning to avoid drying of cells during subsequent culture. Centrifuge at 270 x g, RT for 7 min. Culture at 37 °C in a 5% CO2 incubator. Pseudoislets will complete formation in 5−7 days.
- When harvesting pseudoislets, pre-warm the desired volume of 10% HiFBS CMRL (100 µL per islet), prepare one 50 mL sterile reservoir, one sterile 10 cm Petri dish, and one 8-channel pipette in BSC.
- Remove the 96-well plate from the incubator and place in BSC. Pipette 100 µL per islet of 10% HiFBS CMRL into a reservoir.
- Pipette 100 µL per well of 10% HiFBS CMRL from a reservoir to pseudoislets and pipette up and down 2−3 times gently in the well to lift islets up. Then, aspirate medium in the well containing a pseudoislet and eject into a 10 cm Petri dish. Use of an 8-channel pipette allows transfer of 8 pseudoislets at one time.
- Check the plate under a light microscope to ensure the complete removal of all the pseudoislets. Pseudoislets form firm aggregates and remain aggregated after lifting. The pseudoislets are now ready for downstream experiments.
- Determine the desired number of cells per pseudoislet and the number of pseudoislets to create. Typically, 1,000−3,000 cells are used for each pseudoislet for a 96-well ultra-low attachment plate. For 3,000 cells per pseudoislet, adjust the cell suspension to 1 x 105 cells/mL by resuspending the islet pellet from step 2.10 in 10% HiFBS CMRL so that 30 µL of cell suspension has 3,000 cells. Calculate the total volume of 1 x 105 cells/mL of single cell suspension (mL) required using the following equation:
- Protocol using a 24-well microwell culture plate
- Warm up the anti-adherence rinse solution (Table of Materials) to RT for efficient spheroid formation. Also, pre-warm plain CMRL-1066 and 10% HiFBS CMRL.
- Add 500 µL per well of the anti-adherence rinsing solution to each well of the 24-well microwell culture plate to be used for pseudoislets. Centrifuge at 1,300 x g for 5 min in a swinging-bucket plate centrifuge.
- Observe the plate under a microscope to ensure that air bubbles are removed from microwells. If air bubbles are trapped in microwells, centrifuge at 1,300 x g for 5 min again.
- Aspirate the anti-adherence rinsing solution from the wells in a BSC. Rinse each well with 2 mL of warm plain CMRL-1066 once. Aspirate CMRL-1066. Add 0.5 mL/well of warm 10% HiFBS CMRL to each well planned for use. Wells are now ready for loading dispersed human islet cells prepared in step 2.10.
- Determine the total number of cells needed for each well. One well of 24-well microwell culture plate contains 1,200 microwells and forms 1,200 pseudoislets. 500 cells per pseudoislet x 1200 microwells = 6 x 105 cells. Resuspend single cells from step 2.10 to 6 x 105 cells in 0.8 mL of 10% HiFBS CMRL in a sterile 1.5 mL tube.
NOTE: The maximum volume per well is 2 mL. Volume of cell suspension should not exceed 1.5 mL. The protocol describes steps for creating one well of pseudoislet transduced by lentivirus. Scale up depending on the number of wells to be made for each lentivirus. - For viral transduction, add 125 TU/cell to the single cell suspension. Keep the volume of virus below 0.2 mL. Incubate the cell and virus mixture at 37 °C with occasional gentle mixing for 1 h to allow contact of cells with virus before condensation of cells in step 3.2.8.
NOTE: Use concentrated lentivirus so that the volume of lentivirus added is minimum. Lower number of TU per cell is used for Protocol 2 compared with Protocol 1 as the total volume of medium per cell number is less. However, titer required per cell for efficient gene silencing may differ depending on the lentiviral construct and needs optimization.
CAUTION: Lentivirus is classified as biosafety level 2 and can be integrated into DNA of infected cells. - After 1 h, adjust the total volume of the islet cell and virus mixture to 1 mL by adding 10% HiFBS CMRL. If cells form clump after 1 h incubation, disperse into single cell suspension by gentle and quick pipetting 2−3 times. The pipetting is very important for even distribution of cells across microwells. Transfer cell suspension to one well of the 24-well microwell culture plate.
- Immediately following pipetting, centrifuge at 100 x g for 3 min at RT to capture cells into all microwells. Observe under a microscope to verify that cells are evenly distributed in all microwells.
- Culture the microwell culture plate at 37 °C in a 5% CO2 incubator. Pseudoislets will form in 24−48 h. Pseudoislets can be cultured without medium change for up to 7 days.
- When changing the medium for culture beyond 7 days, replace 50%−75% of medium for each medium change as follows. Slowly remove 0.5−1 mL of medium using a P1000 pipette from each well. Add 0.5−1 mL of fresh 10% HiFBS CMRL slowly by placing a tip to the wall of the well to avoid dislodging pseudoislets from the microwell culture plate.
- To prepare for harvesting pseudoislets, warm up the 10% HiFBS CMRL medium. Pseudoislets tend to float up in serum free CMRL-1066 making it hard to pick them.
- Aspirate 0.5 mL of medium from well using a 1 mL pipette and dispense media forcefully back to plate surface to lift up pseudoislets from the microwell culture plate.
- Gently aspirate dislodged pseudoislets using the 1 mL pipette and transfer islets into a non-tissue culture treated 6-well plate. Pass through a small 37 µm reversible strainer placed on a 15 mL conical tube. Pseudoislets will remain on the filter; any unincorporated single cells will flow through.
NOTE: To avoid loss of smaller size pseudoislets through strainer, the strainer may be omitted. - Dispense 1 mL of 10% HiFBS CMRL across the entire surface of the well to dislodge any remaining pseudoislets, aspirate dislodged pseudoislets, and pass through the strainer. Repeat 3x to ensure complete collection of all pseudoislets from wells.
- Observe the microwell culture plate under an invert microscope to ensure that all pseudoislets are collected. Repeat the wash as in step 3.2.14 if pseudoislets remain.
4. RNA Extraction for Evaluation of Gene Silencing Efficiency
- Pick pseudoislets into the RNase free PBS in a 1.5 mL microcentrifuge tube and centrifuge at 300 x g for 3 min at 4 °C. Remove PBS without disturbing the islet pellet and wash once with PBS followed by centrifugation at 300 x g for 3 min at 4 °C.
- Aspirate most of PBS using a P1000 pipette. Then, change to a P10 pipette to remove the rest of PBS without disturbing the islet pellet.
- Add 0.5 mL of guanidinium thiocyanate RNA extraction reagent (Table of Materials) per tube. Homogenize the pseudoislets using a motor-driven pestle for 2−3 times. The homogenate in the guanidinium thiocyanate RNA extraction reagent can now be stored at -80 °C or processed for RNA purification.
NOTE: 24 of the 3,000 cell-pseudoislets formed in a 96 well plate or 48 of the 500 cell-pseudoislets formed in a microwell culture plate are sufficient to obtain 0.5−1 µg of RNA.
Representative Results
Figure 1 illustrates key steps in the production of pseudoislets using a 96-well ultra-low attachment plate and a microwell culture plate. Figure 2a shows sequential changes in morphology during the formation of pseudoislets from 3 x 103 human islet cells in a 96-well ultra-low attachment plate. Monolayer or loose clumps of cells observed in day 1 changed into solid aggregates with a smooth, round border by day 5 to 7 (Figure 2a). In a microwell culture plate, the formation of solid pseudoislets is usually visible within 4 days (Figure 2b). When 600 cells/microwell were plated in the microwell culture plate, human islet cells were condensed into spheroids of uniform size. It is observed that a microwell culture plate allows the successful formation of pseudoislets from a small number of cells compared with a 96-well ultra-low attachment plate. Typically, over 1,500 cells/pseudoislet are needed for a 96-well ultra-low attachment plate, while 500 cells/pseudoislet are sufficient for a 24-well microwell culture plate. Successfully formed pseudoislets remain as spheroids after recovery from a 96 ultra-low attachment plate or a microwell culture plate and are compatible for downstream applications including static incubation (Figure 3a) and perifusion (Figure 3b). The uniform size of pseudoislets reduces the variation within a test group and allows static incubation using as little as 5 pseudoislets per measurement (Figure 3a). Also, human pseudoislets maintained robust first phase insulin secretion in response to glucose after 7 days of culture when the original human islets cultured for a similar period of time showed blunted first phase glucose-stimulated insulin secretion (Figure 3b)5. The introduction of lentivirus into a single cell suspension ensures the efficient and homogenous transduction of islet cells and achieves highly efficient down-regulation of genes as shown in Figure 3c. All results shown were obtained using human islets from non-diabetic donors.
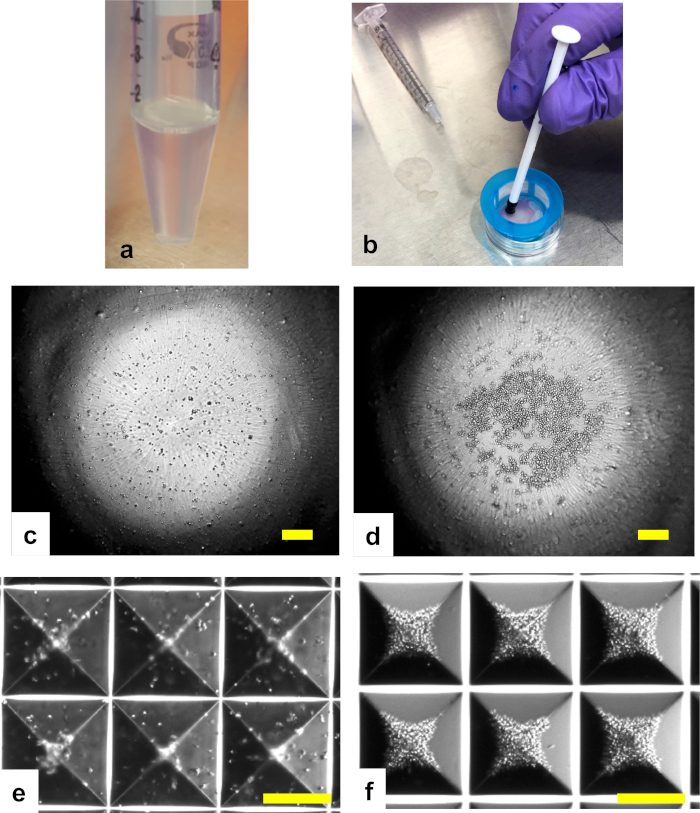
Figure 1: Process of human pseudoislet preparation. (a) The suspension containing 4,000 IEQ of human islets becomes cloudy after digestion by a proteolytic and collagenolytic enzyme mixture and mild pipetting. (b) Human islets after dispersion are passed through a strainer. Undigested islets remaining on top of the strainer are dispersed using a 1 mL syringe plunger. (c,d) Microscope images of the single cell suspension containing 3,000 cells/well in a 96-well ultra-low attachment plate before (c) and after (d) centrifugation. (e,f) Microscope images of the single cell suspension containing 500 cells/microwell in a 24-well microwell culture plate before (e) and after (f) centrifugation. Scale bar = 250 µm. Please click here to view a larger version of this figure.
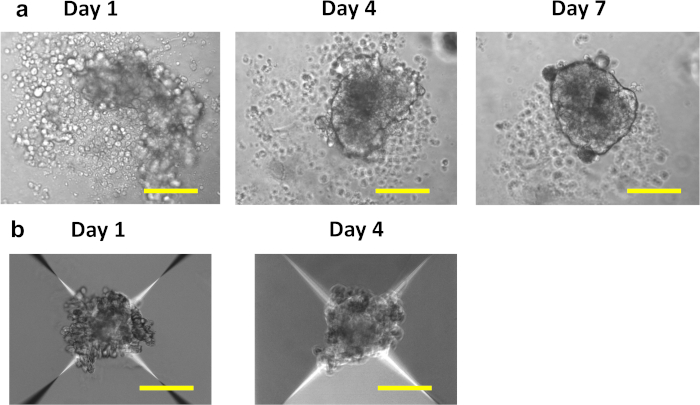
Figure 2: Morphology of human pseudoislets. Sequential changes in morphology of human pseudoislets created (a) in a 96-well ultra-low attachment plate from 3,000 cells and (b) in a 24-well microwell culture plate from 500 cells. Scale bar = 100 µm. Please click here to view a larger version of this figure.
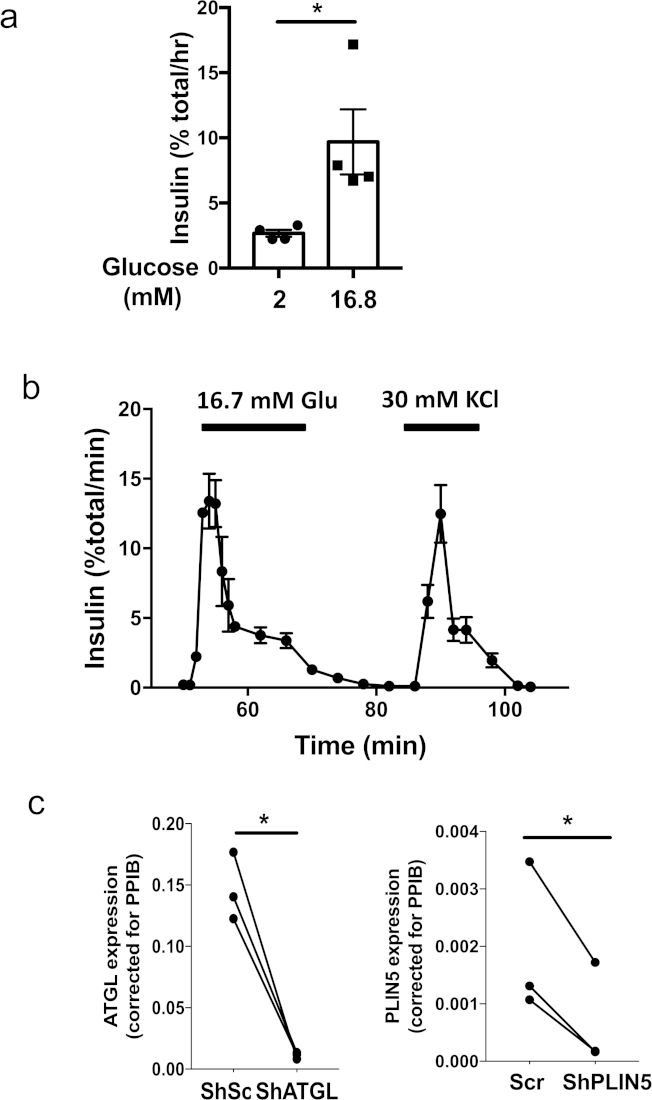
Figure 3: Examples of functional assays using human pseudoislets. (a) Representative static incubation performed using human pseudoislets created in a microwell culture plate from a single donor at the size of 500 human islet cells per pseudoislet. Four sets of 5 pseudoislets were incubated for 1 h in Krebs-ringer bicarbonate buffer supplemented with either 2 mM or 16.8 mM glucose. Each symbol represents insulin secretion from one set of 5 pseudoislets. Mean ± standard error of the mean (SEM) is shown. *, p < 0.05 by student’s t test. Representative results from three donors. (b) Representative perifusion testing insulin secretion from human pseudoislets created in a microwell culture plate in response to 16.7 mM glucose and 30 mM KCl. Method for perifusion was previously published5. Mean ± SEM of insulin secretion from two sets of 40 pseudoislets created in a microwell culture plate from a single donor at the size of 500 human islet cells per pseudoislet plate is shown. Representative data from six donors. (c) Pseudoislets were created with lentivirus carrying shRNA targeting human ATGL (targeting CCTGCCACTCTATGAGCTTAA, left) or PLIN5 (targeting GACAAGCTGGAAGAGAAGCTT, right). Control pseudoislets were transduced with lentivirus expressing scrambled sequence (Scr) previously published5. The mRNA expression of each gene was determined by real time polymerase chain reaction (PCR) as previously published5. Data were expressed using 2–DDCT taking peptidylprolyl Isomerase B (PPIB) as an internal control17. Each dot represents data from each donor for an indicated primer and a data set from the same donor is connected by a line. N = 3 donors. *; p < 0.05 by student’s t test. Please click here to view a larger version of this figure.
Discussion
Here, a detailed protocol to generate human pseudoislets that are transduced by lentivirus using a 96-well ultra-low attachment plate or a microwell culture plate is presented. Pseudoislets have been reported to demonstrate morphology and secretory functions similar to native human islets and can be cultured for prolonged time in vitro5,11,18. Unlike native human islets that show a wide variation in size, pseudoislets are relatively uniform in size, reducing variation between donors and experimental replicates5,11. Downregulation of genes requiring high efficiency of transduction can be performed easily prior to the formation of pseudoislets in single cell suspension. This method avoids the difficulty of viral penetration through layers of cells in intact islets. Thus, this simple, highly efficient, and reproducible protocol for the creation of human pseudoislets has wide applications.
While several different platforms have been reported for the formation of pseudoislets12,14,19,20, both a 96-well ultra-low attachment plate and a microwell culture plate are commercially available, allowing this technique to be adopted by any laboratory. Although the hanging drop method12 also allows the formation of human pseudoislets using common labware, potential limitations include the difficulty in controlling size and reaggregation duration of pseudoislets. These limitations were due to the limited volume per drop of pseudoislet and ongoing evaporation during the 5−7 day culture required for pseudoislet formation. Additionally, it is easier to contain lentivirus with the use of a 96 well ultra-low attachment plate or a microwell culture plate compared with the hanging-drop method.
Several steps within the protocol require close attention. Optimizing the digestion of intact islets with the proteolytic and collagenolytic enzyme mixture is critical since both under- and over-digestion will reduce yield of single cell suspension and subsequently affect aggregation of pseudoislets. During digestion, it is important to closely monitor islets for the disappearance of clumps and the increase in cloudiness as islets dissociate into single cells. It is important to note that the optimal time for digestion varies between islets from different donors. The optimal time is dependent on several factors including medical history and age of each donor, the length of ischemia time, the islet isolation procedure used, islet size, islet purity, islet viability, and shipping conditions. Typically, human islets with viability and purity higher than 80% and within 5 days of isolation are used. Careful and gentle pipetting during dispersion is also important to maintain cell viability and recovery that will ultimately affect cell aggregation and the final size of pseudoislets being formed. When dispensing islet cell suspension to wells (steps 3.1.4 and 3.2.7), gentle and thorough mixing of cells is important to achieve an even distribution of single cells into microwells. If cells form clumps after 1 h incubation with lentivirus, it requires gentle pipetting to break the clumps into single cell suspension prior to the final centrifugation.
We have had similar success in creating human pseudoislets after lentiviral transduction using both a 96-well plate and microwell culture plate. The choice between the two platforms depends on the size and number of pseudoislets desired. A microwell culture plate has small, pyramid shaped bottoms allowing condensation of a smaller number of cells compared with a 96 well round bottom plate. Thus, the number of cells per pseudoislet can be reduced for a microwell culture plate. Also, a single centrifugation step creates all pseudoislets simultaneously in a microwell culture plate while multiple pipetting is required for creating pseudoislets in a 96-well plate. Thus, scaling up the creation of pseudoislets is easier in a microwell culture plate. However, the currently available microwell culture plate does not offer flexibility in the number of pseudoislets being created. Currently, the minimum number of pseudoislets created using a microwell culture plate is 1,200 and can be increased only by the factor of 1,200. Thus, we typically use a 96-well plate for small scale pilot experiments and for an experiment in which small quantity of samples is sufficient such as an insulin secretion assay and RNA extraction for gene expression. We have used pseudoislets from a microwell culture plate for assays that require large numbers of cells such as Western blot, oxygen consumption rate determination by a metabolic analyzer, and triglyceride extraction.
The major limiting factor for the generation of human pseudoislets is the loss of cells during the preparation of single cell suspension. While 1 IEQ of human islet is considered to contain around 2,000 cells, the recovery of single cell suspension is typically 30% or lower due to multiple washing and passing through a strainer. Heterogeneity of islet size also makes it difficult to dissociate all islets simultaneously. While gentle pipetting and the use of the proteolytic and collagenolytic enzyme mixture in the protocol are efforts to combine mechanical and enzymatic forces for maximum recovery of single cells, there still is an inevitable loss of cells. Thus, the application of pseudoislets requires clear justification over studying intact human islets. It also needs to be reminded that insulin secretion from pseudoislets is more robust than islets cultured for the same period of the time but tends to be lower compared with freshly isolated islets5,11.
Although limitations exist, stable and highly efficient gene silencing combined with better preservation of glucose-stimulated insulin secretion for prolonged time in culture enable the assessment of gene function in human islet cells. Additionally, the complex intercellular communication between beta-beta and beta-non beta cells is proposed to have a regulatory role in islet function. However, there is currently limited information regarding intercellular communication within human islets. With increased availability of cell specific markers21, it is feasible to create human pseudoislets with defined cell composition, as recently reported in mouse islet cells22, which will facilitate improved understanding of the cell-cell communication between human islet cells. The recent advancement of imaging of three-dimensional tissues also potentially increases the utility of human pseudoislets as a model to unmask how cellular polarity and intercellular communication are regulated in human islets. Thus, human pseudoislets provide a useful model to dissect the functions of genes of interest and other questions in the field of islet biology.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work was financially supported by National Institutes of Health to Y.I. (R01-DK090490) and American Diabetes Association to Y.I. (1-17-IBS-132). J.A. and Y.I. are supported by the Fraternal Order of Eagles Diabetes Research Center. A.B. is supported by a National Institutes of Health training grant (T32NS45549). Authors utilized human pancreatic islets provided by the NIDDK-funded Integrated Islet Distribution Program (IIDP) at City of Hope (2UC4DK098085).
Materials
| Anti-adherence rinsing solution | Stemcell technologies | 7919 | |
| Biological safety cabinet | Thermo Scientific | 1300 Series Type A2 | |
| cell strainer, 40 micrometer | Corning | 431750 | |
| CMRL-1066 | ThermoFisher | 11530037 | |
| CO2 incubator | Thermo Scientific | Heracell VIOS 160i | |
| conical centrifuge tube, 15 mL | VWR | 89039-666 | |
| conical centrifuge tube, 50 mL | VWR | 89039-658 | |
| fetal bovine serum | ThermoFisher | 26140079 | |
| guanidinium thiocyanate RNA extraction reagent | ThermoFisher | 15596026 | Trizol |
| glutamine | ThermoFisher | 25030164 | |
| Hemocytometer | Marien Feld | Neubauer-Improved Bright line | |
| Human serum albumin | Sigma | A1653 | |
| inverted microscope | Fisher brand | 11-350-119 | |
| microcentrifuge | Beckman Coulter | Microfuge 20 | |
| microcentrifuge tube, 1.5 mL | USA Scientific | 1615-5500 | |
| microwell culture plate | Stemcell technologies | 34411 | Aggrewell 400, 24 well |
| motor-driven pestle | GAMUT | #399X644 | |
| non-tissue culture treated dish, 10 cm | Fisher Scientific | FB0875713 | |
| PBS | ThermoFisher | 14190250 | |
| Penicillin-streptomycin | ThermoFisher | 10378016 | |
| Petri dish, 35 mm | Celltreat | 229638 | |
| pipette, 5 mL | DOT Scientific, | 667205B | |
| pipette, 8-channel | VWR | #613-5253 | |
| pipette, 10 mL | VWR | 667210B | |
| pipette, P10 | Denville | UEZ-P-10 | |
| pipette, P200 | Denville | UEZ-P-200 | |
| pipette, P1000 | Denville | UEZ-P-1000 | |
| proteolytic and collagenolytic enzyme mixture | Sigma | A6965 | Accutase |
| reagent reservoir, 50 mL | VWR | 89094-680 | |
| reversible strainer, 37 micrometer | Stemcell technologies | 27251 | |
| swing bucket plate centrifuge | Beckman Coulter | Allegra X-14R | |
| swing bucket rotor | Beckman Coulter | SX4750A | |
| tuberculin syringe, 1 mL | BD | 309659 | |
| ultra low attachment microplate, 96 well | Corning | 4515 |
References
- Chen, C., Cohrs, C. M., Stertmann, J., Bozsak, R., Speier, S. Human beta cell mass and function in diabetes: Recent advances in knowledge and technologies to understand disease pathogenesis. Molecular Metabolism. 6 (9), 943-957 (2017).
- Hong, H., Jo, J., Sin, S. J. Stable and flexible system for glucose homeostasis. Physiological Review E covering statistical, nonlinear, biological, and soft matter physic. 88 (3), 032711 (2013).
- Cryer, P. E. Minireview: Glucagon in the pathogenesis of hypoglycemia and hyperglycemia in diabetes. Endocrinology. 153 (3), 1039-1048 (2012).
- Arrojo e Drigo, R., et al. New insights into the architecture of the islet of Langerhans: a focused cross-species assessment. Diabetologia. 58 (10), 2218-2228 (2015).
- Harata, M., et al. Delivery of shRNA via lentivirus in human pseudoislets provides a model to test dynamic regulation of insulin secretion and gene function in human islets. Physiological Reports. 6 (20), e13907 (2018).
- Barbu, A. R., Akusjarvi, G., Welsh, N. Adenoviral-mediated transduction of human pancreatic islets: importance of adenoviral genome for cell viability and association with a deficient antiviral response. Endocrinology. 146 (5), 2406-2414 (2005).
- Hughes, A., et al. Gene therapy to improve pancreatic islet transplantation for Type 1 diabetes mellitus. Current Diabetes Reviews. 6 (5), 274-284 (2010).
- Jimenez-Moreno, C. M., et al. A Simple High Efficiency Intra-Islet Transduction Protocol Using Lentiviral Vectors. Current Gene Therapy. 15 (4), 436-446 (2015).
- Bonora, E., et al. Prevalence and correlates of post-prandial hyperglycaemia in a large sample of patients with type 2 diabetes mellitus. Diabetologia. 49 (5), 846-854 (2006).
- Halban, P. A., Powers, S. L., George, K. L., Bonner-Weir, S. Spontaneous reassociation of dispersed adult rat pancreatic islet cells into aggregates with three-dimensional architecture typical of native islets. Diabetes. 36 (7), 783-790 (1987).
- Yu, Y., et al. Bioengineered human pseudoislets form efficiently from donated tissue, compare favourably with native islets in vitro and restore normoglycaemia in mice. Diabetologia. 61 (9), 2016-2029 (2018).
- Zuellig, R. A., et al. Improved physiological properties of gravity-enforced reassembled rat and human pancreatic pseudo-islets. Journal of Tissue Engineering and Regenerative Medicine. 11 (1), 109-120 (2017).
- Caton, D., et al. Lentivirus-mediated transduction of connexin cDNAs shows level- and isoform-specific alterations in insulin secretion of primary pancreatic beta-cells. Journal of Cell Science. 116 (Pt 11), 2285-2294 (2003).
- Arda, H. E., et al. Age-Dependent Pancreatic Gene Regulation Reveals Mechanisms Governing Human beta Cell Function. Cell Metabolism. 23 (5), 909-920 (2016).
- Peiris, H., et al. Discovering human diabetes-risk gene function with genetics and physiological assays. Nature Communications. 9 (1), 3855 (2018).
- Schlimgen, R., et al. Risks Associated With Lentiviral Vector Exposures and Prevention Strategies. Journal of Occupational and Environmental Medicine. 58 (12), 1159-1166 (2016).
- Livak, K. J., Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 25 (4), 402-408 (2001).
- Li, N., et al. Engineering islet for improved performance by optimized reaggregation in alginate gel beads. Biotechnology and Applied Biochemistry. 64 (3), 400-405 (2017).
- Ramachandran, K., Peng, X., Bokvist, K., Stehno-Bittel, L. Assessment of re-aggregated human pancreatic islets for secondary drug screening. British Journal of Pharmacology. 171 (12), 3010-3022 (2014).
- Hilderink, J., et al. Controlled aggregation of primary human pancreatic islet cells leads to glucose-responsive pseudoislets comparable to native islets. Journal of Cellular and Molecular Medicine. 19 (8), 1836-1846 (2015).
- Saunders, D. C., et al. Ectonucleoside Triphosphate Diphosphohydrolase-3 Antibody Targets Adult Human Pancreatic beta Cells for In Vitro and In Vivo Analysis. Cell Metabolism. (18), (2018).
- Reissaus, C. A., Piston, D. W. Reestablishment of Glucose Inhibition of Glucagon Secretion in Small Pseudoislets. Diabetes. 66 (4), 960-969 (2017).
