Le relazioni di scala delle dimensioni del corpo, come l’associazione positiva tra la massa corporea e il tasso metabolico, sono ben note a livello di singolo organismo e sono ora studiate a livelli più elevati di organizzazione1,2,3 . Queste relazioni allometriche sono più spesso funzioni di diritto-potere della forma Y – aMb, dove Y è la variabile di interesse (ad esempio, metabolismo, abbondanza o dimensione della gamma domestica), M è la massa corporea di un singolo o medio individual, b è un coefficiente di scala e a è una costante. Per comodità statistica, i dati Y e M sono spesso trasformati in registro prima dell’analisi, quindi modellati con equazioni lineari del log del modulo (Y) , log (a ) e registro b (M), dove b e log ( a) diventano rispettivamente la pendenza e l’intercettazione del modello lineare.
Lo spettro delle dimensioni è un tipo di relazione allometrica che prevede la densità (D, il numero di individui per unità di superficie) o biomassa (B, la massa sommata di individui per unità di area) in funzione di M (vedere la sezione 4 per ulteriori informazioni sull’uso di stime “normalizzate” di D o B.) Come altre relazioni di ridimensionamento tra M e D o tra M e B, lo spettro delle dimensioni svolge un ruolo centrale nell’ecologia di base e applicata. A livello di popolazione, i biologi spesso interpretano le relazioni D 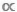 M negative come prova della sopravvivenza dipendente dalla densità o come modelli di capacità di trasporto dell’ecosistema (cioè la “regola di auto-sottilezione”)4, 5. A livello di comunità, le relazioni B M possono essere utilizzate per studiare gli effetti a livello di sistema delle perturbazioni antropogeniche, come la pesca selettiva delle dimensioni6,7. La scala allometrica di D e B con M sono anche al centro dei recenti sforzi per unire l’ecologia della popolazione, della comunità e dell’ecosistema2,8,9.
M negative come prova della sopravvivenza dipendente dalla densità o come modelli di capacità di trasporto dell’ecosistema (cioè la “regola di auto-sottilezione”)4, 5. A livello di comunità, le relazioni B M possono essere utilizzate per studiare gli effetti a livello di sistema delle perturbazioni antropogeniche, come la pesca selettiva delle dimensioni6,7. La scala allometrica di D e B con M sono anche al centro dei recenti sforzi per unire l’ecologia della popolazione, della comunità e dell’ecosistema2,8,9.
Una caratteristica particolarmente importante dello spettro di dimensioni è il fatto che è interamente ataxic9,10. Questo punto è facile da perdere quando si confrontano grafici scatter di dati D M o B M, ma la distinzione tra modelli taxie e atassici è fondamentale. Nei modelli di taxi, un singolo valore M viene utilizzato per rappresentare la massa corporea media di ogni individuo di una data specie o taxa11. Nei modelli atassici, tutti gli individui all’interno di un set di dati sono partizionati tra una serie di intervalli di dimensioni del corpo o contenitori M, indipendentemente dalla loro identità tassonomica12. Quest’ultimo approccio atassico è vantaggioso negli ecosistemi acquatici in cui molti taxa mostrano una crescita indeterminata e sperimentano uno o più cambiamenti ontogenetici nel comportamento alimentare; in questi casi, una singola media M a livello di specie oscurerà il fatto che una specie può ricoprire diversi ruoli funzionali nel corso della sua storia di vita9,13,14.
Qui, presentiamo un protocollo completo per quantificare lo spettro delle dimensioni all’interno di corsi d’acqua guachetabili e fiumi. Il protocollo inizia con i metodi di campionamento sul campo per raccogliere i pesci necessari e i dati macroinvertebrati bentici. I pesci saranno raccolti attraverso un processo di campionamento “esaurimento a tre passaggi”. L’abbondanza sarà quindi stimata dai dati di esaurimento con il metodo15. Nel campionamento di esaurimento, i singoli pesci entro una portata di studio chiusa (cioè, gli individui non possono entrare né uscire dalla portata chiusa) vengono rimossi dalla portata attraverso tre campioni successivi. Pertanto, il numero di pesci rimanenti si esaurirà progressivamente. Da questa tendenza di esaurimento, l’abbondanza totale all’interno della portata dello studio può essere stimata poi convertito in D (nel pesce per m2), utilizzando la superficie nota della portata dello studio. I macroinvertebrati bentici saranno raccolti con campionatori di area fissa standard, quindi identificati e misurati in laboratorio.
Successivamente, i dati combinati di pesci e macroinvertebrati saranno partizionati tra i contenitori di dimensioni. Tradizionalmente, l’ottava o la scala del log2 (cioè gli intervalli di raddoppio) è stata utilizzata per impostare i limiti del contenitore di dimensioni16. Una volta stabilito un elenco di contenitori di dimensioni, il partizionamento dei singoli macroinvertebrati bentici tra i rispettivi contenitori di dimensioni è semplice perché gli invertebrati sono direttamente enumerati come numero di individui per area unitaria. Tuttavia, la stima dell’abbondanza di pesce all’interno dei bidoni di dimensioni è più astratta perché queste stime vengono dedotte dai dati di esaurimento. Vengono quindi fornite istruzioni dettagliate per stimare l’abbondanza di pesce all’interno dei contenitori di dimensioni, indipendentemente dall’identità tassonomica, dai dati del campione di esaurimento.
Infine, la regressione lineare verrà utilizzata per modellare lo spettro delle dimensioni. Questo protocollo è pienamente compatibile con il metodo originale e generale di Kerr e Dickie16 ed identico ai metodi utilizzati da McGarvey e Kirk, 201817 in uno studio degli spettri di dimensioni dei pesci e degli invertebrati nei corsi d’acqua della Virginia Occidentale. Utilizzando questo protocollo, i ricercatori possono assicurarsi che i loro risultati siano direttamente comparabili con altri studi che si basano su Kerr e Dickie16, accelerando così una comprensione ampia e solida delle relazioni di ridimensionamento delle dimensioni del corpo in acqua dolce ecosistemi e i meccanismi che li guidano.
