I mitocondri soddisfano la fornitura chiave di energia e sono un organello compartimentato che contribuisce a processi bioenergetici e metabolici cellulari essenziali come l’anabolismo di nucleotidi, lipidi e amminoacidi, biogenesi del cluster ferro-zolfo e sono implicati nella segnalazione come la morte cellulare controllata 1,2,3 . La bioenergetica mitocondriale attraverso la fosforilazione ossidativa contribuisce a quasi tutti i processi cellulari all’interno della cellula e, di conseguenza, le disfunzioni mitocondriali di origine primaria o secondaria sono associate a un ampio spettro di condizioni di malattia 4,5. La disfunzione mitocondriale comporta non solo alterazioni della struttura o della densità mitocondriale, ma anche della qualità e della regolazione dell’apparato respiratorio6. Questo elemento qualitativo comprende il controllo del substrato, le caratteristiche di accoppiamento, le modifiche post-traduzionali, la dinamica delle cristae e i supercomplessi respiratori 7,8. Pertanto, un’analisi accurata della bioenergetica mitocondriale per approcci sperimentali e diagnostici per valutare il metabolismo energetico della cellula è importante per la salute e la malattia.
La fosforilazione ossidativa mitocondriale (OXPHOS) è una sequenza di reazioni all’interno del sistema respiratorio o del sistema di trasferimento di elettroni (ETS) per la generazione di energia cellulare attraverso l’adenosina trifosfato (ATP)9. La fase multienzimatica per sfruttare l’energia dal flusso di elettroni attraverso i complessi I e II al complesso IV genera un gradiente protonico elettrochimico attraverso la membrana mitocondriale interna, successivamente utilizzato per la fosforilazione dell’adenosina difosfato (ADP) all’ATP tramite il complesso V (F1FO ATP sintasi) (Figura 1A).
In primo luogo, i vettori di due elettroni vengono generati durante il ciclo tricarbossilico (TCA), la glicolisi e l’ossidazione del piruvato: nicotinamide adenina dinucleotide (NADH) e diidroflavina adenina dinucleotide (FADH2). Il NADH viene ossidato nel complesso I (NADH deidrogenasi), durante il quale due elettroni vengono trasferiti al coenzima Q (il chinone viene ridotto a chinolo), mentre i protoni vengono pompati nello spazio intermembrana (IMS). In secondo luogo, il complesso II (Succinato deidrogenasi) ossida FADH2 e alimenta gli elettroni al coenzima Q senza pompare protoni. In terzo luogo, nel complesso III (citocromo c ossidoreduttasi), gli elettroni del coenzima Q vengono trasferiti al citocromo c mentre i protoni vengono pompati nell’IMS. In quarto luogo, il citocromo c trasferisce gli elettroni al complesso IV (citocromo c ossidasi), il complesso finale per pompare protoni e dove l’ossigeno funziona come un accettore di elettroni per assimilare i protoni, formando infine acqua. È questo ossigeno che i mitocondri consumano che può essere misurato da un ossigrafo. Infine, i protoni generati dal complesso I, dal complesso III e dal complesso IV vengono utilizzati per ruotare il complesso V, generando così ATP9.
È importante sottolineare che il trasferimento di elettroni avviene non solo in modo lineare, altrimenti indicato come la catena di trasporto degli elettroni. Invece, gli elettroni possono essere trasferiti al pool di coenzima Q attraverso più vie respiratorie e facilitare il flusso di elettroni convergenti. I substrati nadH e il succinato, ad esempio, possono entrare rispettivamente attraverso il complesso I e il complesso II. Gli elettroni dell’ossidazione degli acidi grassi possono essere donati tramite il complesso flavoproteico che trasferisce elettroni. In effetti, un’analisi completa di OXPHOS richiede un approccio olistico con substrati di carburante appropriati (Figura 1A).
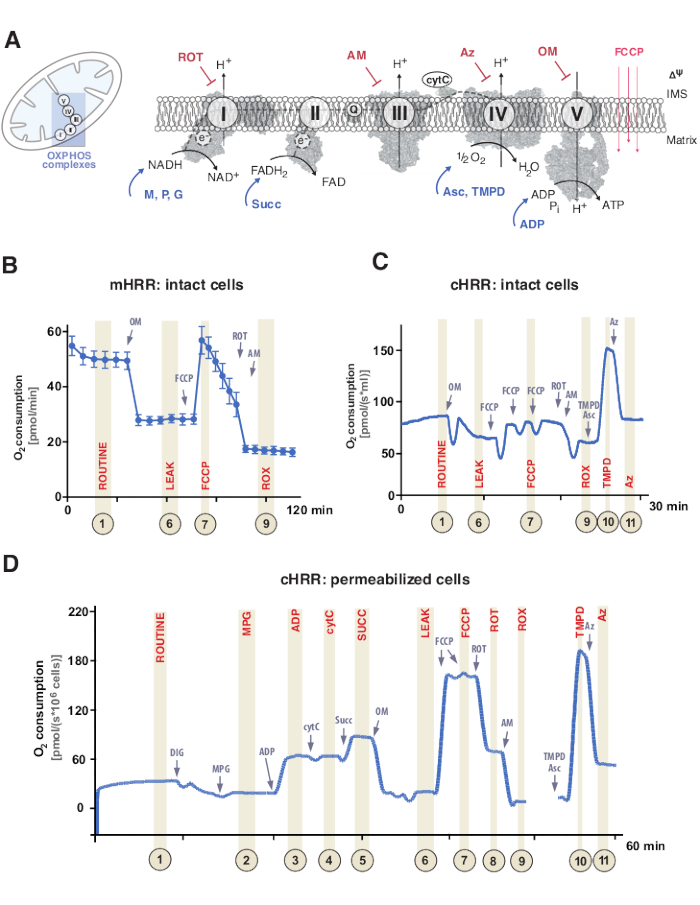
Figura 1: Fosforilazione ossidativa mitocondriale e protocolli specifici di substrato e inibitori. (A) Mitocondrio e schema del sistema di trasferimento di elettroni (CI-CIV) e F1F0 ATP sintasi mitocondriale (CV). Tutte le strutture provengono da PDB. Le figure raffigurano solo substrati e inibitori descritti in questo studio). (B) Traccia campione di flusso di ossigeno in celle HEK293 intatte utilizzando il protocollo standard in un dispositivo mHRR. (C) Traccia campione di flusso di ossigeno in celle HEK293 intatte utilizzando il protocollo standard in un dispositivo cHRR. (D) Traccia campione del flusso di ossigeno nei fibroblasti umani permeabilizzati da un donatore sano con il rispettivo protocollo SUIT. Abbreviazioni: 1 = Respirazione di routine delle cellule intatte; 2 = Stato 2; 3 = Stato 3(I); 4 = Stato 3(I) con cytC; 5 = Stato 3 (I+II); 6 = Perdita(OM); 7 = capacità ETS; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Rotenone, AM = Antimicina, ATP = Adenosina trifosfato, Az = Azide, OM = Oligomicina, FCCP = cianuro di carbonile p-trifluoro-metossifenil-idrazzone; Asc = Ascorbato, TMPD = N,N,N′,N′-tetrametil-p-fenilendiammina, Succ = Succinato, M = Malato, P = Piruvato, ADP = Adenosina difosfato, NAD = Nicotinamide adenina dinucleotide, IMS = Spazio intermembrana, FAD = Flavin adenina dinucleotide. Fare clic qui per visualizzare una versione più grande di questa figura.
L’analisi della capacità di OXPHOS mitocondriale utilizzando HRR è diventata un metodo biochimico strumentale di valore diagnostico non solo per i difetti mitocondriali primari10,11 ma estendendosi a tutti gli altri regni della biologia come il cancro e l’invecchiamento12. HRR consente la determinazione della respirazione cellulare mediante l’analisi della capacità mitocondriale oxphos, che riflette direttamente il deficit del complesso respiratorio mitocondriale individuale o combinato, e indirettamente è associato a disfunzione cellulare e metabolismo energetico alterato9. Metodologicamente, le misurazioni della respirazione vengono eseguite utilizzando cellule, tessuti o mitocondri isolati 11,13,14, con materiale congelato solo parzialmente adatto 15,16. Il tessuto congelato ha dimostrato di avere un ETS intatto con stabilità supercomplessa mantenuta15. Pertanto, a differenza dei tradizionali intermedi TCA, i rispettivi substrati vengono immessi direttamente nell’ETS. Tuttavia, l’accoppiamento tra la sintesi di ETS e ATP viene perso poiché l’integrità della membrana è compromessa da danni da congelamento (formazione di cristalli di ghiaccio).
Gli esperimenti di respirazione si svolgono normalmente a una temperatura fisiologica di 37 °C per gli endotermi in cellule o tessuti non permeabilizzati o permeabilizzati. Mentre il primo considera il contesto metabolico citosolico, il secondo fornisce il contributo energetico dei singoli complessi OXPHOS e dell’ATPasi attraverso l’aggiunta di specifici substrati (e inibitori). La sequenza e la variazione di substrati e inibitori hanno portato allo sviluppo di una vasta gamma di protocolli SUIT17 e saggi18 per affrontare varie questioni scientifiche sulla funzione di OXPHOS (esaminate al punto12). Il protocollo di base della respirazione cellulare valuta quattro diversi stati: i) respirazione di routine – la respirazione in un rispettivo mezzo respiratorio senza alcuna aggiunta di substrati o inibitori che consumano substrati ma endogeni. Questo stato può rivelare OXPHOS generale o difetti respiratori indotti secondariamente causati, ad esempio, da profili metabolici alterati. Successivamente, l’aggiunta dell’oligomicina inibitore dell’ATPasi rivela la permeabilità della membrana mitocondriale interna ai protoni, definita come ii) perdita respiratoria. La successiva titolazione di un protonoforo come il cianuro di carbonile uncoupler p-trifluoro-metossifenil-idrazzone (FCCP) consente di determinare lo stato in cui la capacità ETS è massima in un circuito protonico aperto-transmembrana, definito come iii) respirazione disaccoppiata. È importante sottolineare che uno stato disaccoppiato può verificarsi anche da interventi sperimentali attraverso un eccessivo danno meccanico alle membrane mitocondriali. Al contrario, lo stato non accoppiato si riferisce al disaccoppiamento respiratorio da parte di un meccanismo intrinseco fisiologicamente controllato. Infine, la completa inibizione dell’ETS mediante aggiunta dell’antimicina inibitore del complesso III e dell’inibitore del complesso I rotenone determina il consumo residuo di ossigeno (ROX) da processi non mitocondriali che consumano ossigeno (Figura 1A-C).
La bioenergetica mitocondriale è costituita da cinque distinti stati respiratori19,20. La respirazione dello stato 1 è senza substrati aggiuntivi o ADP, ad eccezione di ciò che è disponibile endogenamente. Dopo l’aggiunta di ADP, ma ancora, nessun substrato, si ottiene la respirazione dello stato 2. Quando vengono aggiunti substrati, consentendo il trasferimento di elettroni e la sintesi di ATP, viene raggiunta la respirazione dello stato 3. In questo stato, la capacità di OXPHOS può essere definita a concentrazioni saturanti di ADP, fosfato inorganico, ossigeno, NADH- e substrati legati al succinato. La respirazione allo stato 4 o la respirazione LEAK può essere definita come uno stato senza ADP o ATP sintasi chimicamente inibite pur avendo substrati sufficienti. Infine, quando tutto l’ossigeno è esaurito (anossico) in un ambiente a camera chiusa, si osserva la respirazione allo stato 5.
Esistono diversi metodi per valutare gli stati energetici cellulari14 con due dispositivi che dominano l’attuale valutazione in tempo reale di OXPHOS attraverso l’analisi del consumo di ossigeno, misurato come funzione della diminuzione dell’ossigeno nel tempo in un sistema a camera chiusa con diversa applicabilità a seconda del modello sperimentale e della domanda di ricerca: il respirometro ad alta risoluzione Oroboros 2k e l’analizzatore di flusso extracellulare Seahorse XF. Entrambi i dispositivi registrano i tassi di consumo di ossigeno come una diminuzione dei picomoli (pmol) di ossigeno (O2) al secondo come valore assoluto all’interno della camera o del pozzo della micropiastra. Il consumo specifico di ossigeno per massa si ottiene normalizzando il rispettivo consumo di ossigeno in una specifica ricetta tampone per numero di cellule (milioni), peso dei tessuti (mg) o quantità di proteine.
L’O2k (Oroboros Instruments) è un sistema chiuso a due camere dotato di un sensore polarografico di ossigeno (abbreviato in respirometro ad alta risoluzione basato su camera: cHRR). Ogni camera sperimentale contiene 2 ml di liquido che viene mantenuto omogeneo da agitatori magnetici. Il sensore polarografico di ossigeno utilizza un approccio amperometrico per misurare l’ossigeno: contiene un catodo d’oro, un anodo di cloruro d’argento / argento e tra una soluzione KCI che crea una cella elettrochimica su cui viene applicata una tensione (0,8 V). L’ossigeno proveniente dal mezzo di analisi si diffonde attraverso una membrana fluorurata di etilene propilene da 25 μm (permeabile A2) e subisce una riduzione al catodo, producendo perossido di idrogeno. All’anodo, l’argento viene ossidato dal perossido di idrogeno, generando una corrente elettrica. Questa corrente elettrica (ampere) è linearmente correlata alla pressione parziale dell’ossigeno. La pressione parziale dell’ossigeno e il fattore di solubilità dell’ossigeno del mezzo di saggio vengono utilizzati per calcolare la concentrazione di ossigeno. Poiché la pressione parziale dell’ossigeno dipende dalla temperatura sperimentale e le misurazioni polarografiche sono sensibili alla temperatura, le fluttuazioni di temperatura richiedono una regolazione precisa (±0,002 °C) da parte di un blocco riscaldante Peltier. La temperatura può essere controllata in un intervallo di 4 °C e 47 °C.
L’analizzatore di flusso extracellulare Seahorse XF (Agilent) è un sistema basato su piastre con formato micropiastra a 24 o 96 pozzetti in cui tre elettrodi di fluorescenza misurano il consumo di ossigeno nel tempo in ciascun pozzo (abbreviato in respirometro ad alta risoluzione basato su micropiastre: mHRR). Un massimo di quattro porte nella cartuccia del test sono disponibili per l’iniezione automatica durante il test. Un test contiene più cicli, ciascuno con tre fasi: 1) miscelazione, 2) attesa e 3) misurazione. Durante la fase di misurazione, le sonde del sensore vengono abbassate nella micropiastra creando una camera temporaneamente chiusa contenente un volume di 7-10 μL per misurare la luce emessa. Questa luce viene emessa da fluorofori incorporati in polimeri sulla punta delle sonde del sensore, che rilevano O2 in base alla tempra a fosforescenza. L’intensità del segnale di fluorescenza è proporzionale a O2 e influenzata dalla temperatura del sensore e del mezzo di saggio. Pertanto, una stima accurata dell’ossigeno richiede un approccio relativo con un pozzo di fondo senza alcun campione. Il ripristino della concentrazione di ossigeno avviene durante la fase di miscelazione quando il sensore si muove su e giù per mescolare il volume sopra la camera temporanea. Ogni ciclo calcola un tasso di consumo di ossigeno. La temperatura può essere controllata in un intervallo di 16 °C e 42 °C.
HRR è il gold standard per valutare la bioenergetica cellulare nelle malattie primarie e associate ai mitocondri e nel metabolismo cellulare generale. In questo studio, vengono forniti protocolli di base per HRR per valutare la funzione di OXPHOS nelle cellule e nei tessuti.
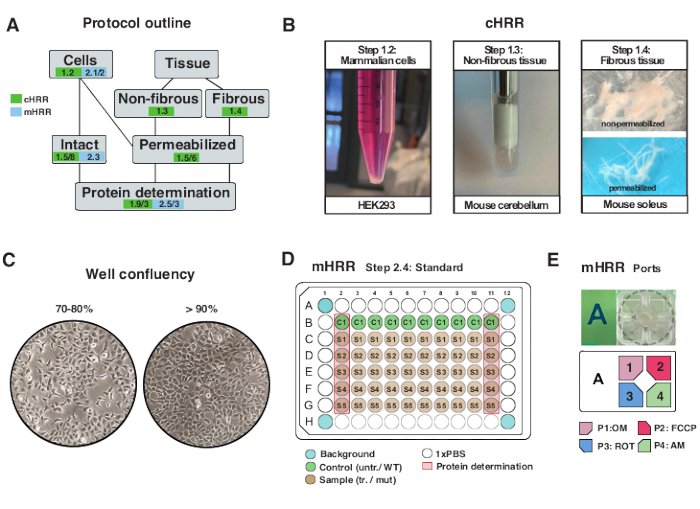
Figura 2: Flusso di lavoro per i preparati cellulari e tissutali per la cHRR e preparazione cellulare per la respirometria mHRR. (A) Schema dei protocolli forniti. (B) Cellule di mammifero (fase 1.2): hek293 pellet pari a 3 x 106 celle (pannello di sinistra). Tessuto non fibroso (fase 1.3): Preparazione del lisato di cervelletto murino in vasaio di teflon da 2 ml (pannello centrale). Permeabilizzazione del muscolo scheletrico indotta da saponina (pannello destro passo 1.4) per la respirometria cHRR. (C) Layout standard di semina delle micropiastre (fase 2.4) e controllo della confluenza per l’analisi delle cellule eucariotiche (HEK293) per la respirometria mHRR. (D, E) Schema di carico della porta di iniezione per la respirometria mHRR (fase 2.4). Fare clic qui per visualizzare una versione più grande di questa figura.
