Un gran numero di eventi di motilità all’interno delle cellule eucariotiche sono mediati dalla funzione delle proteine motorie molecolari. Questi motori si muovono lungo i filamenti citoscheletrici, i filamenti di actina e i microtubuli (MT) e convertono l’energia chimica dell’idrolisi dell’ATP in forze cinetiche e meccaniche necessarie per guidare la motilità biologica all’interno delle cellule. La S. cerevisiae Cin8 basata su MT è una proteina motoria bipolare omotetramerica kinesin-5 che reticola e fa scorrere le MT a parte1. Cin8 svolge funzioni essenziali durante la mitosi, nell’assemblaggio del mandrino 2,3,4 e nell’allungamento del mandrino durante l’anafase 5,6,7. In precedenza, era stato dimostrato che Cin8 è un motore bidirezionale, che commuta la direzionalità in diverse condizioni sperimentali. Ad esempio, in condizioni di elevata resistenza ionica, i singoli motori Cin8 si muovono verso l’estremità negativa degli MT, mentre nei cluster, nei saggi di scorrimento MT multimotore e tra GLI MT antiparallelo, i motori Cin8 si muovono principalmente verso le estremità più degli MT 8,9,10,11,12 . Questi risultati sono stati altamente inaspettati a causa di diversi motivi. In primo luogo, Cin8 porta il suo dominio motorio catalitico all’ammino-terminale e tali motori erano precedentemente ritenuti esclusivamente diretti verso il plus-end, mentre Cin8 ha dimostrato di essere minus-end diretto a livello di singola molecola. In secondo luogo, si credeva che i motori kinesin fossero unidirezionali, diretti meno-end o plus-end, mentre Cin8 si è dimostrato bidirezionale, a seconda delle condizioni sperimentali. Infine, a causa dell’orientamento MT al mandrino mitotico, il ruolo classico dei motori kinesin-5 nella separazione dei poli del mandrino durante l’assemblaggio del mandrino e l’anafase B potrebbe essere spiegato solo dalla loro motilità diretta plus-end sugli MT che reticolano 1,13. A seguito dei primi rapporti sulla bidirezionalità di Cin8, alcuni altri motori kinesin sono stati dimostrati bidirezionali 14,15,16, indicando che la motilità bidirezionale dei motori kinesin potrebbe essere più comune di quanto si credesse in precedenza.
È stato precedentemente riportato che nelle cellule, Cin8 si muove anche in modo bidirezionale8, sostenendo l’idea che la motilità bidirezionale di alcuni motori kinesin-5 sia importante per le loro funzioni intracellulari. Inoltre, poiché i tre motori kinesin-5 che sono stati segnalati come bidirezionali provengono da cellule fungine, un possibile ruolo per la bidirezionalità dei motori kinesin-5 è stato recentemente proposto in tali cellule10. Secondo questo modello, nella mitosi chiusa delle cellule fungine, dove l’involucro nucleare non si rompe durante la mitosi, i motori kinesin-5 forniscono la forza iniziale che separa i poli del mandrino prima dell’assemblaggio del mandrino. Per eseguire questo compito, prima della separazione dei poli del mandrino, i motori kinesin-5 si localizzano vicino ai poli del mandrino, per la loro motilità diretta all’estremità inferiore su singoli MT nucleari. Una volta in questa posizione, i motori kinesin-5 si raggruppano, commutano la direzionalità, catturano e collegano incrociato i NET dai poli del mandrino vicini. Successivamente, i motori kinesin-5 forniscono la separazione iniziale dei poli mediante motilità diretta plus-end sugli MT che reticolano. Con questo modello, sia la motilità diretta minus-end su singoli MT che la motilità diretta plus-end su MT reticolati durante lo scorrimento antiparallelo sono necessarie affinché i motori fungini kinesin-5 svolgano i loro ruoli nell’assemblaggio mandrino 1,13.
L’obiettivo generale del metodo descritto è quello di ottenere kinesin-5 Cin8 fungina di elevata purezza marcata con GFP e di eseguire saggi di motilità a singola molecola (Figura 1) analizzando separatamente la motilità di singole molecole e cluster di Cin8. La separazione tra singole molecole e cluster è importante poiché uno dei fattori che ha dimostrato di influenzare la direzionalità di Cin8 è il suo accumulo in cluster sulle MT10,12. Saggi di motilità alternativi, come i saggi di scorrimento superficiale MT e MT sliding non forniscono informazioni sull’attività delle proteine monomotore17,18. I robusti metodi di analisi e analisi della motilità a singola molecola qui descritti sono stati applicati con successo per caratterizzare diversi aspetti dei motori kinesin-5, Cin8 e Kip1 10,11,12,14,19,20.
Qui viene presentato un protocollo dettagliato per la sovraespressione e la purificazione di Cin8, la polimerizzazione delle MT e il test di motilità a singola molecola. Inoltre, vengono descritte anche le analisi per differenziare tra singole molecole e cluster di Cin8 e per determinare le velocità di singolo motore e cluster mediante l’analisi dello spostamento medio (MD) e dello spostamento quadrato medio (MSD). Questo protocollo ha lo scopo di aiutare i ricercatori a visualizzare tutte le fasi delle procedure e assistere nella risoluzione dei problemi di questo tipo di test.
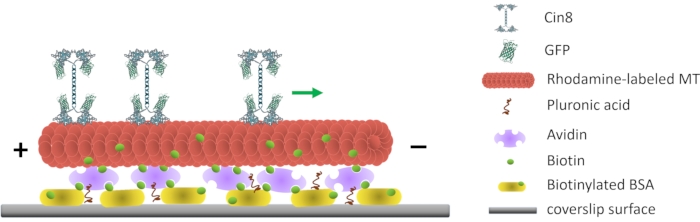
Figura 1: Rappresentazione schematica del saggio di motilità a singola molecola. Le MT fluorescenti biotinilate sono attaccate alla superficie del vetro, rivestite con Avidin che interagisce con il BSA biotinilato attaccato alla superficie. La freccia verde rappresenta la direzione del movimento di singole molecole Cin8 in condizioni di elevata forza ionica. +/- rappresenta la polarità della MT. Fare clic qui per visualizzare una versione più grande di questa figura.
