To show successful transformation using the method above, C. vulgaris was cocultured with either AGL-1 containing the pCAMBIA1302 plasmid or without the plasmid (wild-type and plated on TAP agar supplemented with Hygromycin B and cefotaxime (Figure 1A). The leftmost plate shows the transformed colonies capable of growth on Hygromycin B/cefotaxime plates, and the middle plate shows that wild-type AGL-1 cannot grow on the Hygromycin B/cefotaxime plates. The rightmost plate shows that when no selection is used (cefotaxime only), C. vulgaris transformants and wild-type can grow. Single colonies were streaked onto TAP agar with Hygromycin B and cefotaxime (Figure 1B). Escaped colonies from the rightmost plate (Figure 1B) were inoculated onto TAP liquid with Hygromycin B and cefotaxime (Figure 1C). Due to variable expression levels after random integration of T-DNA, transformants were originally recovered on plates with increasing concentrations of Hygromycin B ranging from 25-70 mg/L. One colony from each plate was grown in liquid TAP media containing the same concentration of Hygromycin B along with the wild-type C. vulgaris at the lowest concentration (Figure 1D). The wild-type C. vulgaris cannot grow in the presence of Hygromycin, while colonies resistant up to 70 mg/L were obtained.
To confirm that the T-DNA cassette was stably integrated into the genome of C. vulgaris, one colony selected from each of the 20, 25, 30, 50, and 70 mg/L Hygromycin B plates were each subcultured by repeated streaking on TAP agar plates with selection two times and then routinely subcultured in TAP media over ten times, named transformant #20, #25, #30, #50, #50, and #70 respectively. Using these cultures, both colony PCR was performed to rule out contamination by pCAMBIA1302 in AGL1, and genomic DNA was extracted to confirm the mGFP5 gene was present in the C. vulgaris genome. In Figure 2A, colony PCR was performed using primers targeting a 5 kbp region outside of the T-DNA left and right borders were able to amplify this region from pCAMBIA1302 plasmid as a positive control, but this region was not present in any of the C. vulgaris samples indicating that pCAMBIA1302 DNA was not present in any of the cultures. The wild-type C. vulgaris sample was used as a negative control, and no amplification was seen.
Genomic DNA was extracted from transformants #25, #50, and #70, and PCR was performed, amplifying a 1763 bp region containing the mgfp5 gene of the T-DNA to confirm integration in the transformants (Figure 2B). pCAMBIA1302 was used as a positive control, and amplification was detected in all of the transformants, indicating that the mgfp5 gene was present in all of the transformants' genomic DNA. The wild-type C. vulgaris was used as a negative control, and no amplification was seen. Figure 2C shows the amplification of a 600 bp fragment of the virulence protein E2 (VirE2) which is found in the tumor-inducing plasmid (Ti plasmid) of the AGl1 strain. The positive control, consisting of the AGl1 strain containing pCAMIA1302, was successfully amplified with primers, indicating the presence A. tumefaciens in the sample. However, all of the C. vulgaris samples tested negative for this region, indicating that repeated subculturing on cefotaxime successfully eliminated contamination of AGL1 from the culture. The wild-type C. vulgaris sample was employed as a negative control, and no amplification was observed. Taken together, these three tests confirm that the T-DNA containing mgpf5 was inserted in the genome of C. vulgaris, and these results were not due to the contamination of AGL1 in the samples nor due to the presence of pCAMBIA1302 in the cells. These results were repeated after 20 and approximately 100 subcultures of the transformants confirming the long-term integration of the T-DNA into the genome.
Finally, the phenotype was confirmed by growing the transformants in TAP media with selection and measuring the fluorescence. This was compared to the wild-type C. vulgaris. Due to the background absorbance/autofluorescence of chlorophyll, the fluorescence was compared by normalizing the data to the wild-type strain. In Figure 3A, it can be observed that there is a noticeable difference in growth between the transformants and the wild-type C. vulgaris. This could potentially be attributed to the multiple random integrations of the T-DNA into the genome, which may be impacting other cellular processes and leading to slower growth. Nonetheless, Figure 3B highlights that despite the lower growth, all of the selected strains exhibit higher fluorescence levels when normalized for cell density. Notably, strain #70 demonstrated significantly higher fluorescence levels than the others, with a p-value of <0.01. This emphasizes the importance of screening numerous colonies to identify transformants that express the desired protein without negatively impacting overall growth behavior.
To confirm the expression of mGFP5-6xHis, SDS-PAGE was used to analyze the crude protein extract and his-tag purified proteins from both C. vulgaris (WT) and C. vulgaris transformant (#50). Subsequently, his-tag protein purification was done using a Ni-NTA resin gravity flow kit following the manufacturer's protocol18. Figure 4 shows the presence of mGFP5-6xHis protein (~30 kDa) in the transformant sample #50 and no protein in the negative control (WT C. vulgaris).
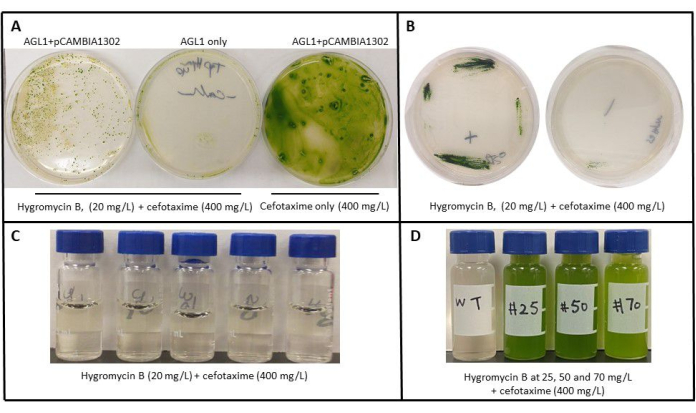
Figure 1: Recovery of C. vulgaris transformants after AMT. (A) After co-cultivation of C. vulgaris, transformants are AGL-1 containing the pCAMBIA plasmid or without the plasmid (AGL-1 only) on selective plates. (B) Re-streaked single colonies selected from Hygromycin B (20 mg/L) plates on selective TAP agar. (C) Escaped colonies lack growth from AGL1 only plate on Hygromycin B (20 mg/L) liquid media. (D) Growth of single colonies #25, #50, and #50 from Hygromycin selection plates grown in liquid TAP media containing Hygromycin and wild-type C. vulgaris that cannot grow in Hygromycin media. Please click here to view a larger version of this figure.
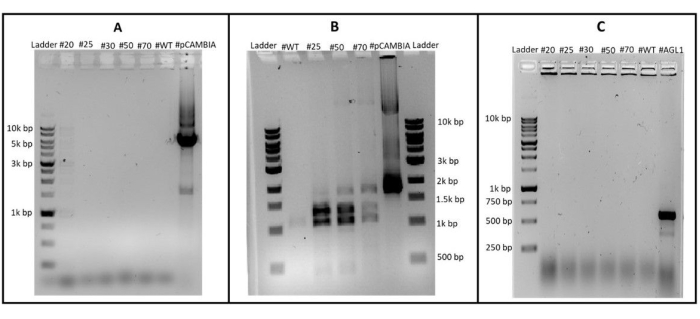
Figure 2: Detection of pCAMBIA1302 contamination. (A) Integration of T-DNA in transformants (B) and detection of A. tumefaciens contamination (C). WT indicates wild-type C. vulgaris, PC is the positive control (pCAMBIA1302) and AGL1 indicates A. tumefaciens AGL1 strain. Please click here to view a larger version of this figure.
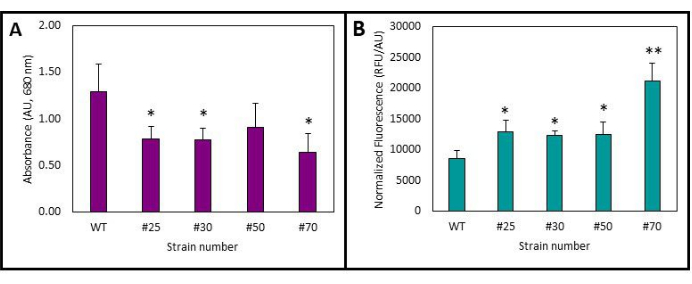
Figure 3: Growth and fluorescence of the transformants at 3 days post-inoculation. (A) Growth as measured by absorbance at 680 nm. (B) Fluorescence normalized to the wild-type strain. Average of 3-4 biological replicates, error bars represent 1 σ. *p < 0.05, **p < 0.01 when compared to the wild-type (WT). Please click here to view a larger version of this figure.
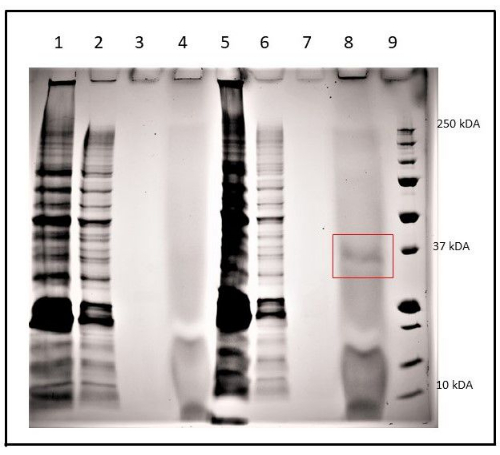
Figure 4: SDS-PAGE analysis of mGFP5-6xHis expression in C. vulgaris (WT) and transformed C. vulgaris sample #50. Extracts were prepared from crude, purified (first wash), purified (diluted), and purified (concentrated) from C. vulgaris (WT), lanes 1-4, respectively. Crude, purified (first wash), purified (eluted), and purified (lyophilized) from C. vulgaris (WT), lanes 5-8, respectively. The red box indicates purified GFP-6xHis protein from transformant #70. Proteins were separated on a polyacrylamide 12% gel. The molecular weights (MWs) of the protein standards (Std.) are shown on the right. Please click here to view a larger version of this figure.
| Name of media/buffer | Composition | ||
| LB | 5 g/L yeast extract, 10 g/L tryptone, 5 g/L NaCl | ||
| TAP | 20 mM Tris base, 1.58 mM K2HPO4, 2.4 mM KH2PO4, 7.0 mM NH4Cl, 0.83 mM MgSO4, 0.34 mM CaCl2, 1 mL/L glacial acetic acid, and 1 mL/L of each F/2 medium trace metals and F/2 vitamins | ||
| F/2 medium trace metals | 22 mg/L ZnSO4·7H2O, 180 mg/L MnCl2·4H2O, 6.3 mg/L Na2MoO4·2H2O, 14 mg/L CoCl2·6H2O, 9.8 mg/L CuSO4·5H2O, 3.15 g/L FeCl3·6H2O, 4.36 g/L Na2EDTA·2H2O | ||
| F/2 medium vitamins | 0.1 mM Vitamin B12 (cyanocobalamin), 25 mg/L Biotin, 335 mg/L Thiamine, 50 mM HEPES Buffer pH 7.8 | ||
| Lysis Buffer | 20 mM Na2HPO4, 300 mM NaCl, pH 7.4 | ||
Table 1: Media and buffer recipes.
