Ökaryotik hücrelerdeki çok sayıda hareketlilik olayı, moleküler motor proteinlerin işlevi tarafından aracılık edilir. Bu motorlar sitoiskelet filamentleri, aktin filamentleri ve mikrotübüller (MTs) boyunca hareket eder ve ATP hidrolizinin kimyasal enerjisini, hücreler içindeki biyolojik hareketliliği yönlendirmek için gereken kinetik ve mekanik kuvvetlere dönüştürür. MT bazlı S. cerevisiae Cin8, iş mili MT’lerini çapraz bağlayan ve kaydıran bipolar, homotetramerik kinesin-5 motor proteinidir1. Cin8, mitoz sırasında, iş mili montajı 2,3,4’te ve anafaz 5,6,7 sırasında iş mili uzamasında temel işlevleri yerine getirir. Daha önce, Cin8’in farklı deneysel koşullar altında yönselliği değiştiren çift yönlü bir motor olduğu gösterilmişti. Örneğin, yüksek iyonik mukavemet koşulları altında, tek Cin8 motorları MT’lerin eksi ucuna doğru hareket ederken, kümelerde, çok motorlu MT kayma testlerinde ve antiparalel MT’ler arasında, Cin8 motorları esas olarak MTs 8,9,10,11,12’nin artı uçlarına doğru hareket eder. . Bu bulgular birkaç nedenden dolayı oldukça beklenmedikti. İlk olarak, Cin8 katalitik motor alanını amino terminusta taşır ve bu tür motorların daha önce sadece artı uçlu yönlendirildiğine inanılıyordu, oysa Cin8’in eksi uçlu tek molekül seviyesinde yönlendirildiği gösterildi. İkincisi, kinesin motorlarının eksi uçlu veya artı uçlu tek yönlü olduğuna inanılıyordu, oysa Cin8’in deneysel koşullara bağlı olarak çift yönlü olduğu gösterildi. Son olarak, mitotik mildeki MT oryantasyonu nedeniyle, kinezin-5 motorlarının iş mili montajı ve anafaz B sırasında iş mili direklerinin ayrılmasındaki klasik rolü, yalnızca 1,13’ü çapraz bağladıkları MT’ler üzerindeki artı uçlu yönlendirilmiş hareketlilikleri ile açıklanabilir. Cin8’in çift yönlülüğü hakkındaki ilk raporları takiben, diğer birkaç kinesin motorunun çift yönlü 14,15,16 olduğu gösterildi, bu da kinesin motorlarının çift yönlü hareketliliğinin daha önce inanıldığından daha yaygın olabileceğini gösteriyor.
Daha önce hücrelerde, Cin8’in çift yönlü bir şekilde hareket ettiğibildirilmiştir 8, bazı kinezin-5 motorlarının çift yönlü hareketliliğinin hücre içi fonksiyonları için önemli olduğu fikrini desteklemektedir. Ek olarak, çift yönlü olduğu bildirilen üç kinesin-5 motoru mantar hücrelerinden olduğundan, bu tür hücrelerde yakın zamanda kinesin-5 motorlarının çift yönlülüğü için olası bir rol önerilmiştir10. Bu modele göre, nükleer zarfın mitoz sırasında parçalanmadığı mantar hücrelerinin kapalı mitozunda, kinesin-5 motorları, iş mili montajından önce iş mili kutuplarını ayıran ilk kuvveti sağlar. Bu görevi yerine getirmek için, iş mili kutup ayrımından önce, kinesin-5 motorları, tek nükleer MT’ler üzerindeki eksi uçlu yönlendirilmiş hareketlilikleriyle iş mili kutuplarının yakınında lokalize olur. Bu konuma geldiğinde, kinesin-5 motorları kümelenir, yönselliği değiştirir, komşu mil direklerinden MT’leri yakalar ve çapraz bağlar. Daha sonra, kinesin-5 motorları, kutupların çapraz bağladıkları MT’lerde artı uçlu yönlendirilmiş hareketlilik ile ilk ayrımını sağlar. Bu modele göre, mantar kinezin-5 motorlarının iş mili düzeneği 1,13’teki rollerini yerine getirmesi için hem tek MT’lerde eksi uçlu yönlendirilmiş hareketlilik hem de çapraz bağlı MT’lerde artı uçlu yönlendirilmiş hareketlilik gereklidir.
Tarif edilen yöntemin genel amacı, yüksek saflıkta fungal GFP etiketli kinesin-5 Cin8 elde etmek ve tek moleküllerin ve Cin8 kümelerinin motilitesini ayrı ayrı analiz ederken tek moleküllü motilitesi testleri (Şekil 1) yapmaktır. Tek moleküller ve kümeler arasındaki ayrım önemlidir, çünkü Cin8’in yönlülüğünü etkilediği gösterilen faktörlerden biri, MTs10,12 üzerindeki kümelerde birikmesidir. MT yüzey kayma ve MT kayma testleri gibi alternatif hareketlilik testleri, tek motor proteinlerinin aktivitesi hakkında bilgi sağlamaz17,18. Burada açıklanan sağlam tek moleküllü hareketlilik testi ve analiz yöntemleri, kinezin-5 motorlarının, Cin8 ve Kip1 10,11,12,14,19,20’nin farklı yönlerini karakterize etmek için başarıyla uygulanmıştır.
Burada, Cin8 aşırı ekspresyonu ve saflaştırılması, MTs’lerin polimerizasyonu ve tek moleküllü motilitesi testi için ayrıntılı bir protokol sunulmaktadır. Ayrıca, tek moleküller ve Cin8 kümeleri arasında ayrım yapmak ve ortalama yer değiştirme (MD) ve ortalama kare yer değiştirme (MSD) analizi ile tek motor ve küme hızlarını belirlemek için yapılan analizler de açıklanmaktadır. Bu protokol, araştırmacıların prosedürlerin tüm adımlarını görselleştirmelerine ve bu tür tahlillerde sorun gidermeye yardımcı olmalarına yardımcı olmayı amaçlamaktadır.
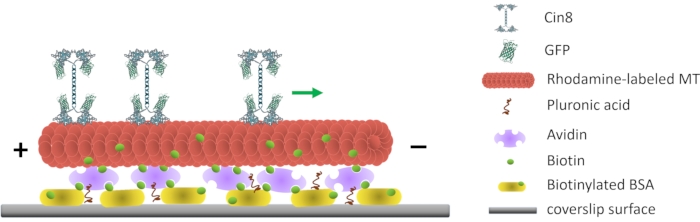
Şekil 1: Tek moleküllü motilitesi testinin şematik gösterimi. Biyotinile floresan MTs, yüzeye bağlı biyotinillenmiş BSA ile etkileşime giren Avidin ile kaplanmış cam yüzeye tutturulur. Yeşil ok, yüksek iyonik mukavemet koşulları altında tek Cin8 moleküllerinin hareket yönünü temsil eder. +/- MT’nin polaritesini temsil eder. Bu rakamın daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
