A simple approach to determine the spatial spread of iontophoresis is to retract the iontophoretic pipette stepwise from the dendrite, while keeping the ejected glutamate constant. We found that the spatial extent of a micro-iontophoretic stimulation had a diameter of approximately 12 μm (Figure 1 showing radius). How deep in the tissue the iontophoresis can be used depends on the rigidity of the pipette. However, the iontophoretic pipettes needed for experiments in slices (Figure 2), which are used here, are not limiting the depth of penetration. Rather the optical system and the decreasing resolution in the depth of the brain slice is the limiting factor. For a good quality recording it is crucial that no transmitter is leaking out of the pipette. In the case of glutamate, a leakage can be identified if there is a sudden depolarisation when the dendrite is approached with the pipette tip or if the baseline suddenly becomes unstable (Figure 5). After establishing a stable recording, it is possible to evoke EPSPs of defined amplitudes, dendritic spikes or action potentials with this technique on any location throughout the dendritic tree (Figure 6). By applying glutamate with fast micro-iontophoresis, the properties of EPSPs, their propagation and summation, simultaneously at different even distant locations, can be investigated (Figure 7). GABAergic events can be evoked alone or in addition to glutamate with a second pipette by filling the iontophoretic pipette with a highly concentrated GABA solution (see: Protocol section 2) and a positive eject current. There is a simple protocol to confirm the GABAergic nature of the events and to make GABAergic events easier to detect, if not using a high driving force internal solution: Inject negative currents (approx. 1 sec; the amplitude depends on the input resistance of the cell) resulting in hyperpolarising voltage steps, starting at around -100 mV and then increase in 5 mV steps (Figure 8). At very negative potentials the GABAergic events are easier to detect, due to the higher driving force. And if the signals reverse around the calculated Cl– reversal potential for the solutions, it is very likely that they are GABAergic in nature (Figure 8). With an additional GABA micro-iontophoretic pipette, it is possible to investigate, for example, the effect of GABAergic inhibition on glutamatergic events, like dendritic sodium/calcium spikes, by varying the relative timing of dendritic spike and IPSP (Figure 9), the relative location of both events, or their amplitudes. (All animal experiments were conducted in accordance with the guidelines of the Animal Care and Use Committee of the University of Bonn, the German Center for Neurodegenerative Diseases and the state Northrhine-Westfalia.)
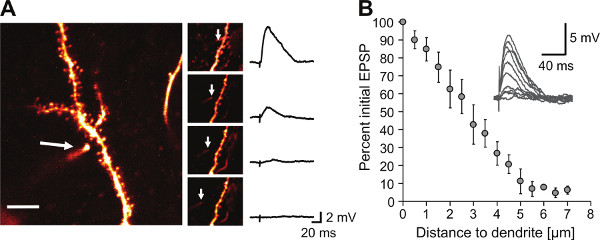
Figure 1. Spatial extent of iontophoretically ejected glutamate determined by pipette retraction. A) Maximum intensity projection of a two-photon image of a CA1 pyramidal cell dendrite filled with 100 μM Alexa 594 and the iontophoretic pipette in the initial position (scale bar 8 μm). Insets show retraction of the iontophoretic pipette and the corresponding iontophoretic EPSP recorded at the soma. Arrow indicates the position of the iontophoretic pipette tip. B) Distance dependence of EPSP amplitudes relative to the initial position of the iontophoretic pipette tip (≤1 μm away from dendrite, n = 6 branches). With this we could estimate the radius of the glutamate spread by systematically retracting the pipette. Inset shows representative example traces. Error bars represent mean ± SEM. (Adapted from Müller et al. 20129, reprinted with permission from Elsevier).
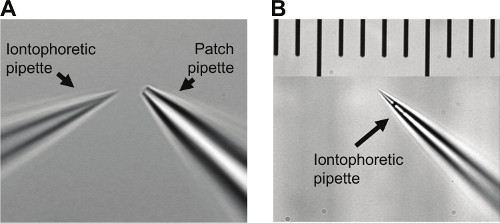
Figure 2. How an iontophoretic pipette should look like. A) Infrared CCD image of an iontophoretic pipette compared to a small 5.5 MΩ patch pipette using a 60X objective. B) Iontophoretic pipette with scale using a 60X objective. Calibration: smallest = 10 μm.

Figure 3. Capacitance compensation of the iontophoretic pipette using the build in test-pulse (10 nA, 10 msec, npi electronic, Tamm, Germany). It is important to compensate the capacitance correctly to monitor the right pipette resistance and to ensure fast and accurate neurotransmitter application.
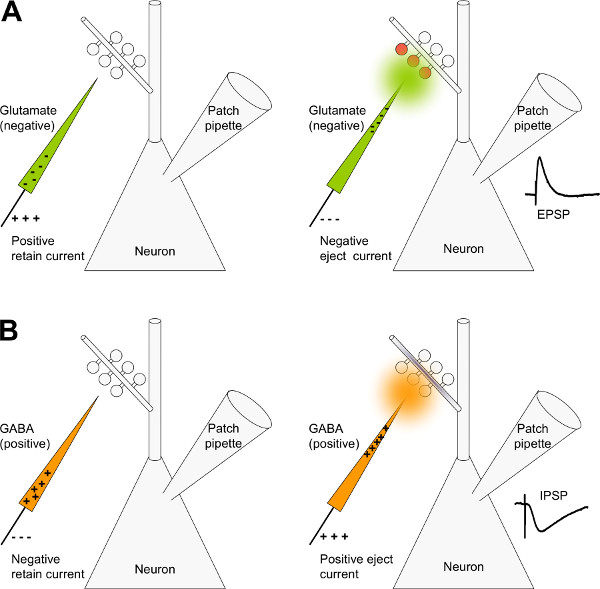
Figure 4. Principles of micro-iontophoresis. A) Schematic drawing of a CA1 pyramidal neuron in whole cell patch-clamp configuration and an iontophoretic pipette filled with glutamate. Glutamate is negatively charged, therefore a positive current applied to the pipette will keep it from leaking out of the pipette: The retain current is positive (left panel). To eject glutamate from the pipette, apply a negative current (right panel). In this way glutamate is forced out of the pipette and can evoke excitatory events in the postsynaptic cell. B) Schematic drawing of CA1 pyramidal neuron in whole cell patch-clamp configuration and an iontophoretic pipette filled with GABA. GABA is positively charged at a low pH. Therefore, a negative current will keep it from leaking out of the pipette (left panel). To eject GABA apply a positive current (right panel). In this way GABA comes out of the pipette and can evoke inhibitory events in the postsynaptic cell.
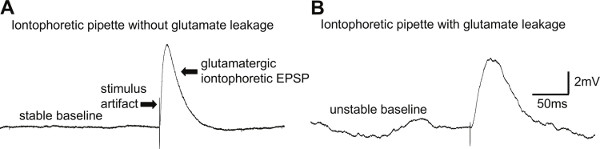
Figure 5. Good and bad iontophoretic pipettes. A) Representative example of an iontophoretic glutamatergic EPSP generated on a dendritic branch of a CA1 pyramidal neuron. B) Representative example of an iontophoretic dendritic spike generated a dendritic branch in the CA1 area of the hippocampus with ongoing mild glutamate leakage from the pipette tip.
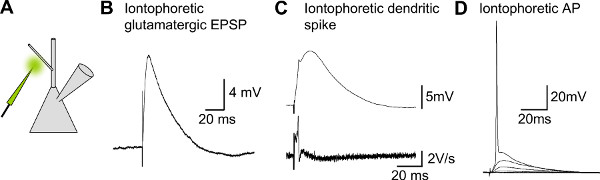
Figure 6. Representative results for single glutamate micro-iontophoresis. A) Schematic drawing of a patched CA1 pyramidal neuron and an iontophoretic pipette filled with glutamate. B) EPSP evoked in a CA1 pyramidal neuron with glutamate iontophoresis. C) Dendritic Na+/Ca2+spike evoked on a proximal dendrite of a CA1 pyramidal neuron, lower trace shows the slope of the voltage trace, peak indicates the peak slope of the dendritic spike. D) When increasing the current applied to the iontophoretic the amplitude of the EPSP will increase until it crosses the action potential threshold.
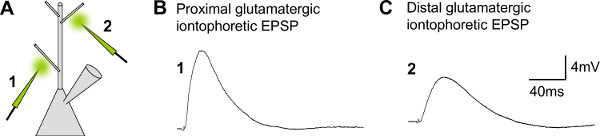
Figure 7. Representative results for double glutamate micro-iontophoresis at different locations on the dendritic tree. A) Schematic drawing of a patched CA1 pyramidal neuron and two iontophoretic pipettes filled with glutamate, which are positioned on a proximal, and a distal dendrite, respectively. B) Iontophoretic EPSP evoked on a proximal dendrite of a CA1 pyramidal neuron (1). C) Iontophoretic EPSP evoked at a distal dendrite (2).
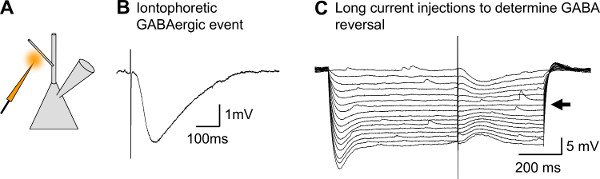
Figure 8. Representative results for GABA micro-iontophoresis. A) Schematic drawing of a patched CA1 pyramidal neuron and an iontophoretic pipette filled with GABA. B) Iontophoretic IPSP evoked on a proximal dendrite of a CA1 pyramidal neuron. C) Long current injection of systematically changing amplitudes to a CA1 pyramidal neuron via the patch pipette, to determine the reversal potential of the evoked event (current injections from -400 pA to 0 pA). Artifact indicates time-point of GABA iontophoresis. The evoked event indicates a reversal potential of approximately -70 mV (arrow).
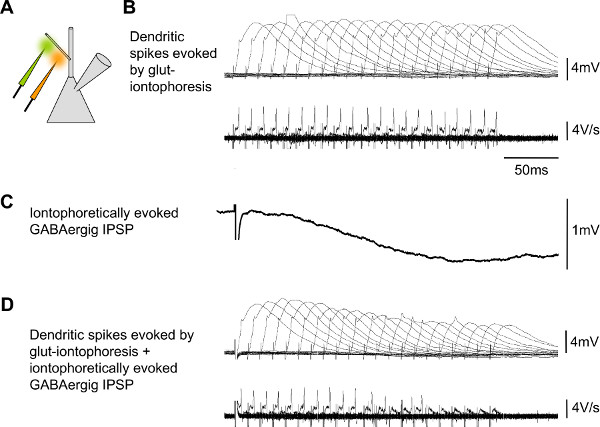
Figure 9. Representative results of simultaneous glutamate and GABA iontophoresis to investigate integration of inhibition and excitation. A) Schematic drawing of a patched pyramidal neuron and one pipette filled with glutamate (green) and on with GABA (orange). B) Iontophoretically evoked dendritic spikes alone, in subsequent sweeps, lower traces show dV/dt, peaks indicate dendritic spikes. C) Iontophoretically evoked IPSP alone. D) Both, glutamatergic and GABAergic events together.
| Location specificity | Transmitter specificity | Toxicity / side effects | Presynaptic stimulation | Long-term experiment | Costs/complexity | |
| Micro- iontophoresis | ++ | +++ | ++ | – | ++ | ++ |
| 2-photon uncaging | +++ | +++ | + | – | + | + |
| Synaptic stimulation | – | – | +++ | +++ | +++ | +++ |
Table 1. Comparison of different techniques. Advantages and disadvantages of techniques to stimulate neurons according to different criteria (- = bad, + = not optimal, ++ = good, +++ = best).
| Patch pipettes | Iontophoretic pipettes | |||
| Pre-pulls P (A) Single pull | Last Pull P(B) | Pre-pulls P(A) Single Pull | Last Pull P(B) | |
| Heat H | 700 | 480 | 510 | 600 |
| Force Pre Pull F(TH) | 018 | 035 | 018 | 008 |
| Distance Threshold s(TH) | 017 | 012 | 025 | 015 |
| Delay Heatstop t(H) | 050 | 030 | 050 | 030 |
| Distance Heatstop s(H) | 030 | 000 | 030 | 000 |
| Delay F(F1) | 000 | 136 | 000 | 050 |
| Force Pull 1 F1 | 000 | 065 | 200 | 400 |
| Distance Pull 2 s(F2) | 000 | 005 | 000 | 001 |
| Force Pull 2 F2 | 000 | 080 | 000 | 095 |
| Adjust (AD) | 121 | 000 | 121 | 000 |
Table 2. Exemplarily puller protocol. For a horizontal puller (DMZ-Universal Puller, Zeitz-Instruments GmbH, Martinsried, Germany), using GB150F-8P filaments (Science Products, Hofheim, Germany) for both patch and iontophoretic pipettes.
