Originally, mESCs are maintained on an MEF feeder in mESC medium with FBS and other supplements (Figures 1A and 2A). CM was collected from mESCs in Reduced Serum Media without a feeder layer, FBS, or other supplements (Figures 1B and 2B). This culture condition allows us to collect mESC-specific conditioned medium without potential contamination by the factors from the feeder, FBS, or other supplements. The control medium was collected under the same culture conditions, without mESCs.
mESCs show different morphologies between the two culture media: i) normal mESC culture conditions (Figure 2A) and ii) serum- and feeder-free culture conditions (Figure 2B). The mESC colonies grew on an MEF layer and demonstrated an oval and shiny appearance under the normal mESC culture conditions (Figure 2A). On the contrary, the mESCs in the serum- and feeder-free culture conditions showed a flattened and irregular morphology (Figure 2B).
The functional characterization of mESC-CM was achieved by senescence-associated methods, such as SA β-gal assay (Figure 3A), cell cycle analysis (Figure 3B), and qPCR (Figure 3C). Treatment of senescent HDFs with mESC-CM decreased the number of positive SA β-gal-positive cells, which is an indicator of cellular senescence (Figure 3A). Cell cycle analysis revealed that mESC-CM treatment dramatically increased the number of cells in the S and G2/M phase, whereas it reduced the number of cells in the G0/G1 phase (Figure 3B). In addition, mESC-CM treatment decreased the senescence-associated gene expression levels (namely, p53, p21, and p16) and the senescence-associated secretory phenotype (SASP) expression levels (IL-6).
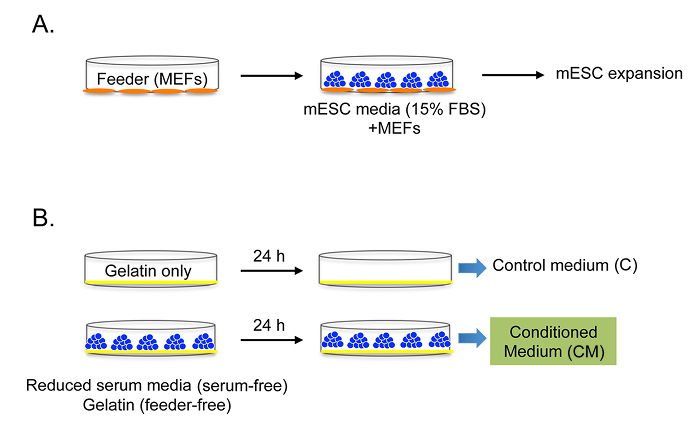
Figure 1: Preparation and optimization of mESC-CM. Experimental strategy for the preparation and optimization of serum-free and feeder-free CM. (A) Normal mESC culture condition and (B) serum- and feeder-free mESC-CM culture condition. C: control medium without FBS and MEF; CM: conditioned medium without FBS and MEF. Modified with permission from Bae et al.15. Please click here to view a larger version of this figure.
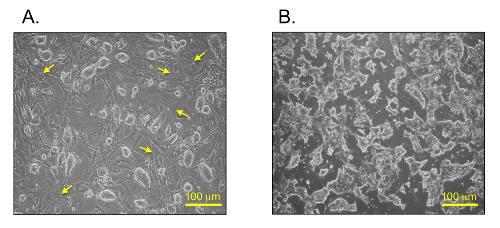
Figure 2: Bright field images of mESCs. mESCs under (A) normal conditions and (B) serum- and feeder-free conditions. Yellow arrows indicate feeder cell (MEFs) in normal mESC culture conditions. Scale bars = 100 µm. Please click here to view a larger version of this figure.
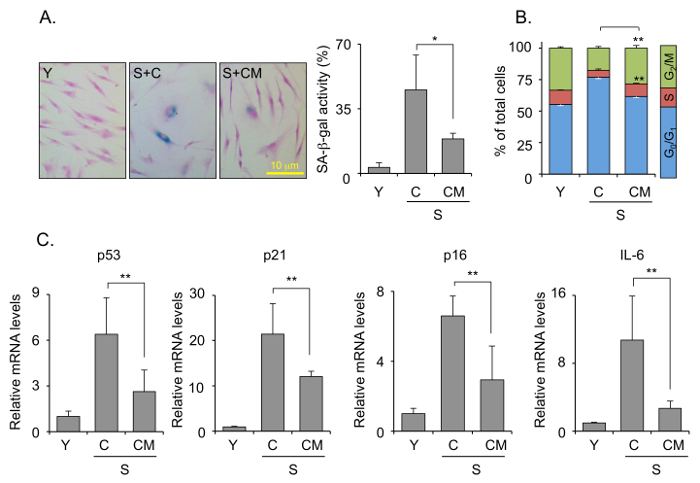
Figure 3: Characterization of the anti-aging effect of mESC-CM. (A) SA β-gal activity staining and the percentage of SA β-gal-positive cells. (B) Cell cycle analysis by flow cytometry. (C) Expression levels of senescence-associated gene expression levels (p53, p21, and p16) and senescence-associated secretory phenotype (SASP) expression levels (IL-6) by qRT-PCR. Values are the mean ± SD. Figures are representative of three independent experiments. Statistically-significant differences between groups were identified by one-way ANOVA and Tukey's post-hoc test. *p <0.05, **p <0.01. Y = non-senescent cells; S: senescent cells; C: control medium without FBS and MEF; CM: conditioned medium without FBS and MEF. Scale bars = 10 µm. Modified with permission from Bae et al.15. Please click here to view a larger version of this figure.
