Thermogenetic screen.
Abnormally increased appetite causes elevated food intake, regardless of physiological needs. We utilized this scheme to design a high-throughput behavioral screen to obtain genetic handles of neurons related to hunger and satiated states (Figure 1). The screen yielded Taotie-Gal424. When the Taotie-Gal4 neurons were forcibly activated at 30 °C, the Taotie>TrpA1 flies ingested larger quantities of food than the controls (Figure 2, Figure 3).
Visually scoring the food consumption.
Because some neurotransmitters and neuropeptides have been indicated in the regulation of feeding behavior, we have tested the corresponding GAL4 lines, including NPF20, sNPF39, octopamine21, dopamine27,40, serotonin41,42, AKH35, and Dilp27,8,35. To quantify the feeding response, we visually inspected and scored flies with various amounts of detectable dye in the gut (Figure 2A). After activation of Taotie neurons, about 58% of Taotie>dTrpA1 flies exhibited strong feeding behaviors, and 23% of these showed mild feeding behaviors (Figure 2B). In contrast, only marginal feeding behaviors were observed in flies with other neurons activated via dTrpA1 (Figure 2B).
Colorimetric quantification of food intake.
To quantify food intake with high precision and objectivity, we measured the absorbance of fly extraction at a wavelength specific to the added dye in the food2,27,28. To correlate an absorbance value with the volume of food-intake, a standard curve was obtained by measuring the absorbance of the sample solutions (the same buffer for homogenizing the flies, PBST) mixed with different amounts of dyes). As our results demonstrate, acute activated Taotie-GAL4 neurons by TRPA1 dramatically increased food intake compared with genetic control and the temperature control during the same test period (Figure 3B), suggesting that the Taotie-GAL4 labeled neurons participate in regulation of food intake of adult Drosophila.
Optogenetic activation to promote food intake in Drosophila.
We used UAS-CsChrimson to activate Taotie neurons by illumination with an orange light36 (Figure 4A, 4B). When Taotie>CsChrimson flies were stimulated by LED lights at 607 nm, they ingested significantly more food than controls (Figure 4C). Furthermore, the amounts of ingested food correlated well with the intensities of stimulation light (Figure 4D). Thus, besides thermogenetics with dTrpA1, optogenetic activation with CsChrimson in Taotie neurons also promotes feeding motivation in satiated flies.
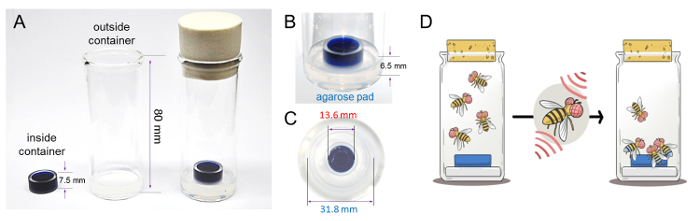
Figure 1. A feeding paradigm for analyzing appetite in adult flies. (A) The feeding chamber contains an inside container filled with dye-colored food and an outside container padded with 1% agarose. From left to right, the inside container, the outside container, and the complete feeding chamber. (B) The inside container sits on top of the agarose pad at the bottom of the outside container. (C) Top view of the feeding chamber. (D) A schematic showing the "appetite screen" experiment. Normal satiated flies rarely ate food (left vial), while forcibly activating certain feeding control neurons caused satiated flies to ingest additional food, thus exhibiting dye-colored food in the abdomen (right vial). Please click here to view a larger version of this figure.
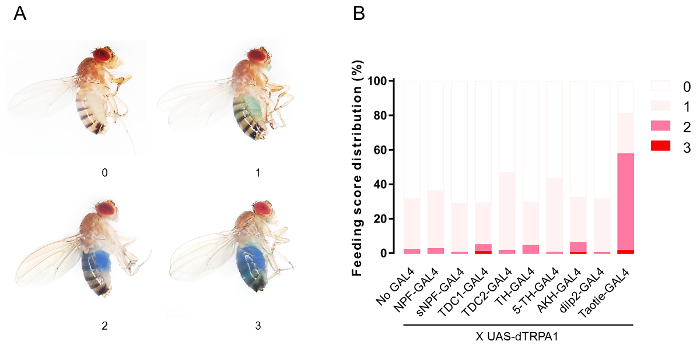
Figure 2. Visual estimation of food ingestion. (A) Exemplar images to show the criteria for scoring food content in the abdomen. (B) The distribution of feeding scores in flies with indicated neurons being activated by dTrpA1 at 30 °C. No-Gal4: UAS-dTrpA1 (genetic control); NPF-GAL4: neuropeptide F positive neurons; sNPF-GAL4: short neuropeptide F positive neurons; TDC1-GAL4 and TDC2-GAL4: octopamine neurons; TH-GAL4: dopamine neurons; 5-HT: serotonin neurons; AKH: adipokinetic hormone positive neurons; dip2: insulin-like peptide 2 positive neurons; Taotie-GAL4: Taotie-GAL4 labels neurons (n = 120 flies per condition). Please click here to view a larger version of this figure.
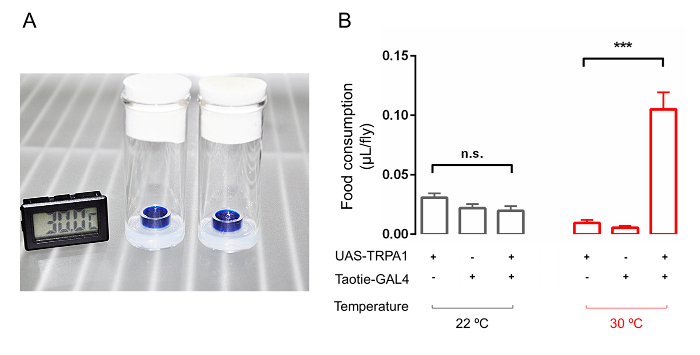
Figure 3. Thermogenetic-activation of Taotie neurons increases food consumption in adult flies. (A) An image shows the setup of the thermogenetic activation experiment. The feeding test was performed in an incubator at 30 °C. (B) Food consumption by colorimetric quantification in satiated flies tested at either 30 °C or 22 °C for 1 h. The amount of food consumption of a single fly was calculated from that of 20 flies in one feeding chamber (n = 6). n.s. indicates not significant (p >0.05); ***p <0.001. One-way ANOVA followed with Tukey's post hoc test was used to analyze multiple comparisons. Error bars indicate mean ± SEM. Please click here to view a larger version of this figure.
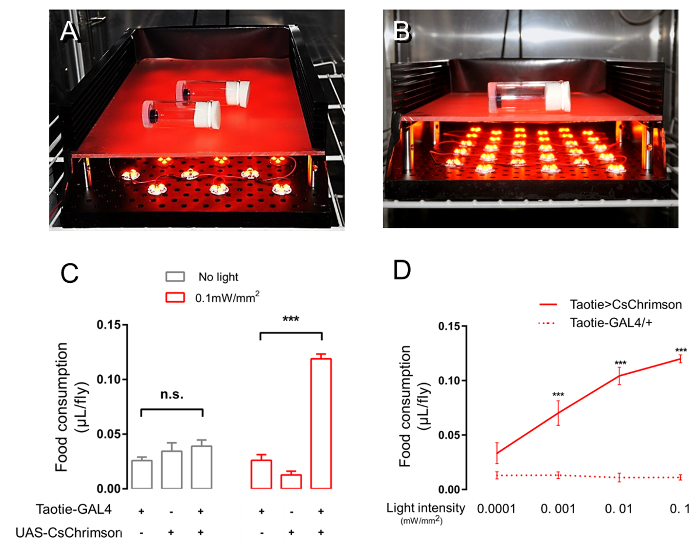
Figure 4. Optogenetic-activation of Taotie neurons increases the amount of food intake in an intensity-dependent manner. (A,B) Two views of the setup for optogenetic activation. The feeding chambers were laid sideways on a supporting plate and illuminated from the bottom by an array of orange LEDs. The front side of the illumination box was removed to show the inside LEDs. The behavioral experiments were performed inside of an incubator at 25 °C, 60% RH. (C) Food consumption of satiated flies of indicated genotypes when tested under optogenetic illumination or in the dark for 1 h (n = 6). (D) Food ingestion by satiated Taotie>CsChrimson flies was correlated with the illumination intensity (n = 6). n.s. indicates not significant (p >0.05); ***p <0.001, One-way ANOVA followed with Tukey's post hoc test was used to analyze multiple comparisons, Student's t-test for two group comparisons. Error bars indicate mean ± SEM. Please click here to view a larger version of this figure.
