The protocol for the enrichment of LDL from chicken egg yolk is illustrated in Figure 1. This process begins by diluting whole egg yolk with saline and separating the egg yolk solids referred to as granules from the soluble or plasma fraction containing the LDLs (Figure 1)33. The LDL content of the plasma fraction is further enriched by precipitation of the ~ 30-40 kDa β-livetins (Figure 2)33. The presence of protein bands at 140, 80, 65, 60 and 15 kDa correlate with the apoproteins of LDLs (Figure 2)33,39. Treatment with triclosan inhibits growth of S. aureus in fatty acid-free media32. We have previously demonstrated that supplementing cultures with egg yolk plasma or purified human LDLs as exogenous fatty acid sources overcomes triclosan-induced growth inhibition (Figure 3)32. Similarly, supplementation of triclosan-treated cultures with enriched egg yolk LDL restores growth (Figure 3). Further, addition of egg yolk LDLs support the growth of a previously characterized S. aureus fatty acid auxotroph (Figure 4)32. For the most accurate mass spectrometry-based profiling of S. aureus incorporation of exogenous fatty acids, it is important to limit the presence of free fatty acids in the growth medium. The free fatty acid composition of 1% tryptone broth and chicken egg yolk LDLs diluted in tryptone broth was determined by employing flow injection high-resolution/accurate mass spectrometry and found minimal quantities of free fatty acid (Figure 5). The same untargeted mass spectrometry analysis was performed to determine the fatty acid composition of S. aureus phospholipids after exposure to chicken egg yolk LDLs. Orthogonal partial least-squares discriminant analysis (OPLS-DA)40 of abundant S. aureus membrane phospholipids demonstrated clear class separation of untreated and chicken egg yolk LDL-treated conditions, as shown in the OPLS-DA scores plot (Figure 6A). The OPLS-DA loadings plot indicated numerous phosphatidylglycerol species as important variables in the PLS-DA model. Notably, phospholipids containing unsaturated fatty acids, a molecular marker of exogenous fatty acid incorporation, are enriched in the LDL supplemented cultures compared to cells incubated in the absence of LDLs (Figure 6B). Previous studies have found that chicken egg yolks are a rich source of unsaturated fatty acids with oleic acid (18:1) being the most abundant41,42. In agreement with these observations, we found oleic acid to be the most common unsaturated fatty acid utilized for phospholipid synthesis when S. aureus cultures were supplemented with chicken egg yolk LDLs (Figure 6C). Table 1 further illustrates that the fatty acid profiles of membrane phospholipids are altered when S. aureus is grown in the presence of egg yolk LDL.
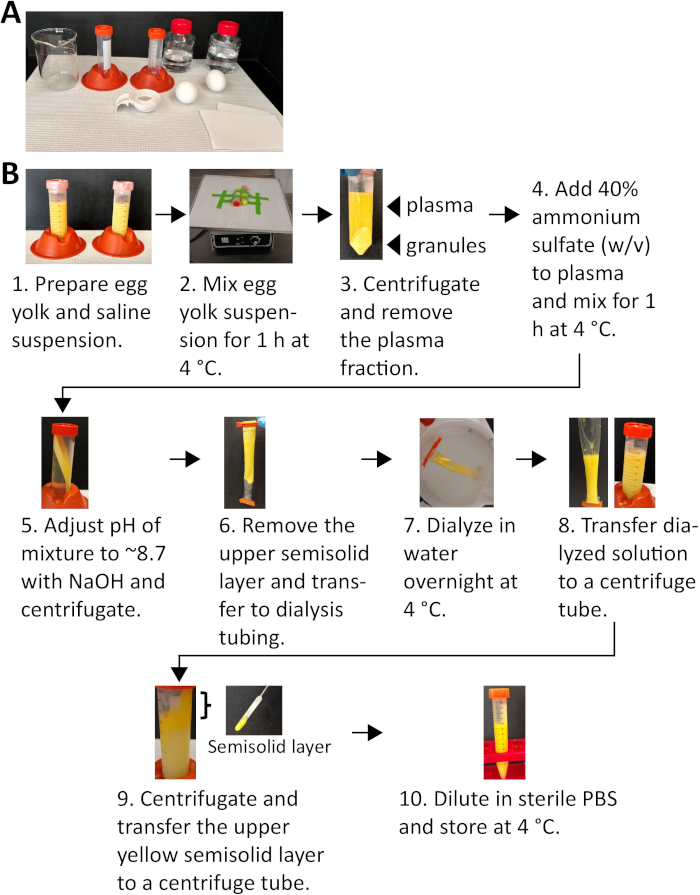
Figure 1: An illustration of LDL enrichment from chicken egg yolk utilizing centrifugation and ammonia sulfate precipitation. (A) The reagents necessary for the enrichment of LDL from chicken egg yolk. (B) The flow chart depicts the significant steps of the LDL enrichment process. Please click here to view a larger version of this figure.

Figure 2: Protein profile of chicken egg yolk prior to and after enrichment for LDL. Protein lysates were prepared using RIPA buffer. Protein lysate (15 µg) was loaded into an 8% acrylamide SDS-PAGE gel. Gels were stained overnight with Bio Rad Protein reagent. The molecular weights in kDa of LDL associated proteins are denoted along the right side of the image. M: protein marker, Y: chicken egg yolk, and LDL: chicken egg yolk LDL enrichment Please click here to view a larger version of this figure.

Figure 3: Egg yolk-derived LDLs protect S. aureus from triclosan-induced FASII inhibition. The growth of S. aureus was monitored over time via measurement of OD600 in 1% tryptone broth in the following conditions: 1% tryptone broth (TB), 1 µM triclosan (TCS), 1 µM triclosan with 1% egg yolk plasma (TCS + EYP), 5% egg yolk LDL (LDL), or 1 µM triclosan with 5% egg yolk LDL (TCS + LDL). The mean from three independent experiments is shown. Error bars represent the standard deviation of the mean. Please click here to view a larger version of this figure.
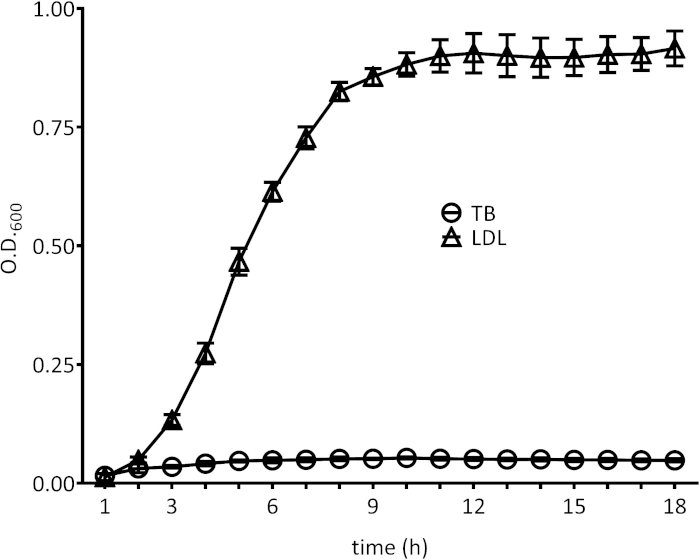
Figure 4: Growth of a S. aureus fatty acid auxotroph is supported by egg yolk-derived LDL. The growth of a fatty acid auxotroph in 1% tryptone broth (TB) with or without 5% egg yolk LDL (LDL) supplementation was monitored over time via measurement of OD600. The mean from three independent experiments is shown. Error bars represent the standard deviation of the mean. Please click here to view a larger version of this figure.

Figure 5: Free fatty acid content measured in 1% tryptone broth or chicken egg yolk LDL. Free fatty acids were detected by flow injection high-resolution/accurate mass spectrometry and tandem mass spectrometry. Normalized numbers of ions per mg of protein was determined for 1% tryptone broth and 1% tryptone broth supplemented with 5% chicken egg yolk LDLs. Please click here to view a larger version of this figure.

Figure 6: Chicken egg yolk low-density lipoproteins are a reservoir of exogenous fatty acids for synthesis of S. aureus phosphatidylglycerol. (A) Scores plot of orthogonal partial least-squares discriminant analysis of chicken egg yolk LDL-treated and untreated S. aureus membrane phospholipids identified using high resolution/accurate mass spectrometry. (B) Percentage of unsaturated phosphatidylglycerol (UPG) compared to total membrane PG of S. aureus grown in the absence (WT) or presence (WT + LDL) of chicken egg yolk LDLs. (C) Unsaturated fatty acid (UFA) profile of membrane PG of S. aureus grown without (WT) or with (WT + LDL) chicken egg yolk LDLs graphed as a percentage of the amount of total PG fatty acids. Please click here to view a larger version of this figure.
| WT cultured in tryptone broth | WT cultured in tryptone broth supplemented with LDLs | |||||
| Phosphatidyl glycerol (TC:TDB)a | Normalized ion abundance/mg of protein | SD | Fatty acidsc | Normalized ion abundance/mg of protein | SD | Fatty Acidsc |
| 24:0 | 0 | 0 | NDb | 0.052031116 | 0.02677 | ND |
| 26:0 | 0 | 0 | ND | 0.009539117 | 0.00362 | ND |
| 28:0 | 0.127937113 | 0.04528 | 15:0_13:0 | 0.167643281 | 0.02392 | 15:0_13:0 |
| 28:1 | 0.006765427 | 0.00157 | ND | 0.002776821 | 0.00372 | 15:0_13:1 |
| 30:0 | 8.680180809 | 2.68375 | 15:0_15:0 | 14.04873592 | 2.4531 | 15:0_15:0 |
| 30:1 | 0 | 0 | ND | 0.010152161 | 0.00449 | 15:1_15:0, 13:1_17:0 |
| 31:0 | 4.150511117 | 1.31658 | 16:0_15:0, 14:0_17:0 | 10.17590926 | 1.88431 | 16:0_15:0, 14:0_17:0, 18:0_13:0 |
| 31:1 | 0.016156004 | 0.01216 | 13:1_15:0, 12:1_19:0 | 0.473478683 | 0.09063 | 13:1_15:0, 18:1_13:0, 12:1_19:0 |
| 32:0 | 29.29259262 | 8.82993 | 15:0_17:0 | 48.24342037 | 8.95664 | 15:0_17:0, 16:0_16:0 |
| 32:1 | 0.02074815 | 0.00941 | ND | 0.307044942 | 0.07305 | 18:1_14:0, 16:1_16:0 |
| 33:0 | 9.000460122 | 2.78194 | 18:0_15:0, 16:0_17:0 | 15.4531776 | 2.98171 | 18:0_15:0, 16:0_17:0 |
| 33:1 | 0.162934812 | 0.04796 | ND | 2.921832928 | 0.30851 | 18:1_15:0 |
| 33:2 | 0 | 0 | ND | 0.167492702 | 0.03211 | 18:1_15:1, 18:2_15:0 |
| 34:0 | 12.3064043 | 3.70242 | 19:0_15:0, 17:0_17:0 | 18.40129157 | 3.21385 | 19:0_15:0, 17:0_17:0 |
| 34:1 | 0 | 0 | ND | 1.423605186 | 0.20066 | 18:1_16:0 |
| 34:2 | 0.000470922 | 0.00082 | ND | 0.156133734 | 0.03929 | 18:2_16:0 |
| 35:0 | 5.727462455 | 1.74583 | 20:0_15:0, 18:0_17:0 | 7.771538992 | 1.28515 | 20:0_15:0, 16:0_19:0, 18:0_17:0 |
| 35:1 | 0.17337586 | 0.05727 | 20:1_15:0 | 0.772202525 | 0.08526 | 20:1_15:0, 18:1_17:0 |
| 35:2 | 0 | 0 | ND | 0.038758757 | 0.01481 | 18:2_17:0, 18:1_17:1 |
| 36:0 | 0.671004303 | 0.2116 | 21:0_15:0, 19:0_17:0 | 0.967295024 | 0.2572 | 21:0_15:0, 20:0_16:0, 19:0_17:0, 22:0_14:0 |
| 36:2 | 0 | 0 | ND | 0.495485065 | 0.04473 | 18:1_18:1, 18:2_18:0 |
| 36:3 | 0 | 0 | ND | 0.059268233 | 0.02291 | 18:2_18:1, 20:3_16:0, 20:2_16:1 |
| 37:0 | 0.060466411 | 0.01961 | 22:0_15:0, 20:0_17:0 | 0.114526894 | 0.01852 | 22:0_15:0, 20:0_17:0, 18:0_19:0 |
| 38:2 | 0 | 0 | ND | 0.079469521 | 0.02872 | 18:2_20:0, 16:1_20:1 |
| aDetected as [M-H]– ions. TC, total chain length; TDB, total number of double bonds. | ||||||
| bND, not determined | ||||||
| cFatty acids are listed in order of isomer abundance. An underscore between fatty acid designations indicates that each fatty acid may be present in either the SN1 or SN2 position, as tandem mass spectrometry alone cannot rule out the possibility that lipid species exist as a mixture of positional isomers. | ||||||
Table 1: Fatty acid profile of S. aureus cultured in the presence of chicken egg yolk LDLs. We used an unbiased lipidomic analysis utilizing high-resolution/accurate MS and MS/MS to determine the fatty acid profile of S. aureus PG. S. aureus was incubated in the presence or absence of chicken egg yolk LDLs, and the PG profile of these cells was compared to that of cells cultured in 1% tryptone broth.
