Body weight changes
A one-way ANOVA did not show any differences in changes in body weight between experimental groups for the 21 days of the diving-for-food task. From days 2 to 21, there were changes in body weight for all 3 groups (p<0.01, Table 1).
Sucrose preference test
At the start of the experiment (day 0), there was no difference in the percent of sucrose preference between the experimental group of rats induced with anhedonia (85.6% ± 18.6), the experimental group treated with antidepressant therapy (85.1% ± 18.8), and the control group (85.7% ± 9.9). On day 35, compared to the control group (84.13% ± 12.3), there was a significantly lower percent sucrose preference in the experimental group (62.69% ± 17.7, p<0.01) and in the experimental group with treatment (68.48% ± 13.9, p<0.01, Figure 3A). There were not yet any differences between the experimental group and the experimental group with treatment. On day 62, the experimental rats had a lower percent sucrose preference (68% ± 15) than the control group (78.5% ± 16) and the experimental group with treatment (77% ± 16, p<0.05, Figure 3B). There were no differences between the treatment group and the control group at this time. Data is presented as percent sucrose preference ± standard deviation.
Diving-for-food task
The social activity of rats in a situation of restricted access to food is illustrated in Figure 4. Rats in the experimental group demonstrated an increase in frequency of entries into the tunnel (113% ± 3.7, p<0.01, Figure 4A), diving for food (141% ± 7, p<0.01, Figure 4B), food obtained by carrying (168% ± 12, p<0.01, Figure 4C), time spent in separate cages (123% ± 7.9, p<0.01, Figure 4D), and food obtained by attack (232% ± 26, p<0.01, Figure 4E) compared to the experimental group with treatment (44% ± 7, 53% ± 6, 54% ± 5, 55% ± 4.7, 67% ± 3.4, respectively). The differences between the experimental group of rats and the experimental rats treated with antidepressants were statistically greater than the difference between the experimental group and the control group in all 5 parameters of the diving-for-food test (p<0.05). Data is presented as an average percentage compared to controls ± standard error of the mean.
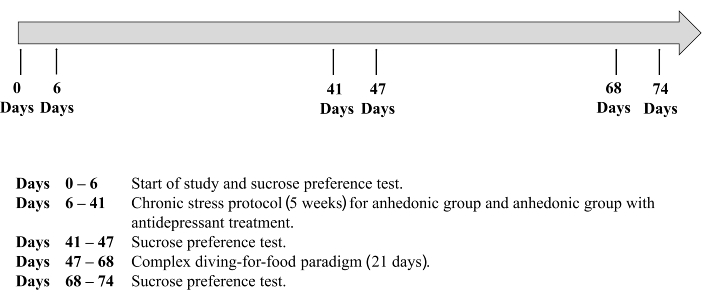
Figure 1. A timeline of the experimental protocol. Please click here to view a larger version of this figure.
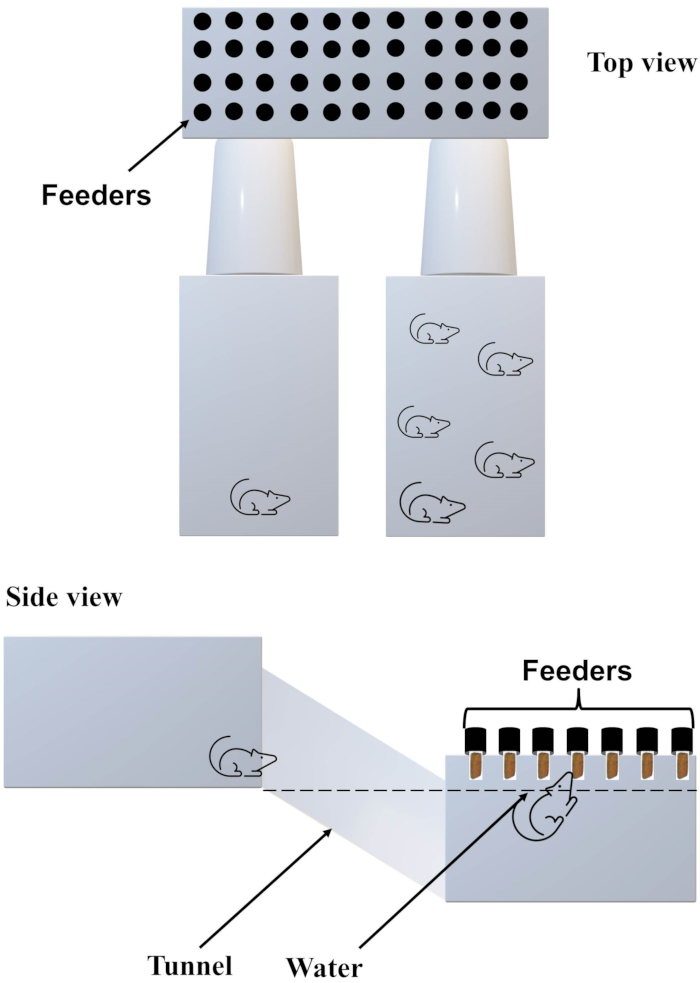
Figure 2. Illustration of the diving-for-food apparatus. Please click here to view a larger version of this figure.
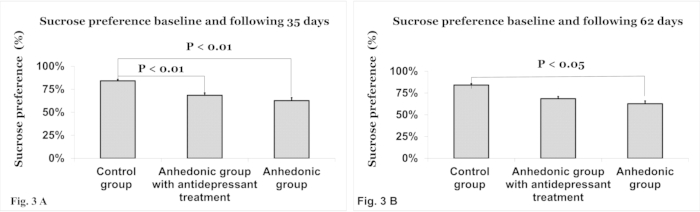
Figure 3. The sucrose preference test (A) after 35 days and (B) after 62 days. There was no difference in sucrose consumption at the beginning of the experiment. (A) By day 35 of the experiment, the anhedonic group (p<0.01) and the anhedonic group treated with antidepressant therapy (p<0.01) had a significantly lower percent sucrose preference than the control group. (B) On day 62, the rats induced with anhedonia had a lower percent sucrose preference compared to both the control and the anhedonic group treated with antidepressant treatment (p<0.05). Please click here to view a larger version of this figure.
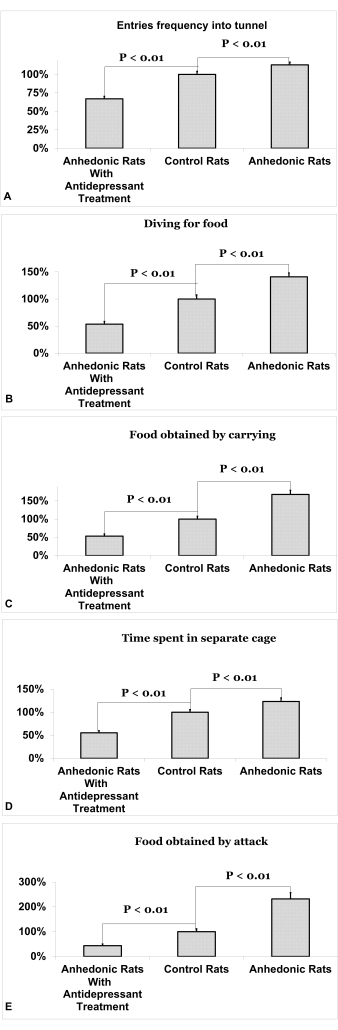
Figure 4. Social activity of rats in a situation of restricted access to food. (A) Frequency of entries into the tunnel. (B) Diving for food. (C) Food obtained by carrying. (D) Time spent in separate cages. (E) Food obtained by attack. Data is presented as an average percentage compared to mean control values + standard error of the mean. Please click here to view a larger version of this figure.
| Change in rats’ body weight | |||||||||||||||||||||
| Days | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 |
| Control Group | |||||||||||||||||||||
| AVER. | 0 | -0.02 | -0.04 | -0.04 | -0.06 | -0.06 | -0.07 | -0.08 | -0.09 | -0.1 | -0.11 | -0.12 | -0.12 | -0.13 | -0.14 | -0.15 | -0.16 | -0.18 | -0.19 | -0.2 | -0.21 |
| SD | 0 | 0.01 | 0.01 | 0.02 | 0.01 | 0.02 | 0.02 | 0.01 | 0.01 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.03 |
| Experimental Group with Treatment | |||||||||||||||||||||
| AVER. | 0 | -0.02 | -0.03 | -0.04 | -0.05 | -0.06 | -0.07 | -0.08 | -0.09 | -0.1 | -0.1 | -0.12 | -0.12 | -0.14 | -0.14 | -0.15 | -0.16 | -0.17 | -0.18 | -0.19 | -0.2 |
| SD | 0 | 0.02 | 0.02 | 0.02 | 0.02 | 0.03 | 0.03 | 0.02 | 0.03 | 0.03 | 0.03 | 0.02 | 0.03 | 0.03 | 0.03 | 0.04 | 0.03 | 0.03 | 0.03 | 0.02 | 0.02 |
| Experimental Group | |||||||||||||||||||||
| AVER. | 0 | -0.01 | -0.03 | -0.04 | -0.05 | -0.05 | -0.07 | -0.07 | -0.08 | -0.1 | -0.11 | -0.12 | -0.13 | -0.14 | -0.15 | -0.15 | -0.17 | -0.18 | -0.19 | -0.2 | -0.21 |
| SD | 0 | 0.01 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.02 | 0.02 | 0.02 |
Table 1. Changes in body weight (as a percentage) during the diving-for-food task. There were no differences between the 3 experimental groups for changes in body weight during the 21 days of the task. From days 2 to 21, there was an overall effect between days expressed as a change in body weight (p<0.01).
