A complete understanding of biological principles requires the study of many diverse organisms spanning the Tree of Life. Although a broad range of organisms were described through the end of the 19th century, by the middle of the 20th century experimental studies became restricted to a handful of less than a dozen model organisms. With the advent of the genomic era and the goal to sequence genomes from all species in the Tree of Life1, we are now in a position to expand the types of organisms being used for laboratory experiments and to reap the advantage of their diversity. Such expansion of emerging model organisms for experiments has a prerequisite of being able to maintain them in the laboratory. Here, protocols are described for rearing one such emerging new/old model organism.
The majority of animal life on Earth is accounted for by four super-radiations of Insects2. Within the Insects, there are about 158,000 species of Diptera (true flies)3, with about 3000 species in the family Sciaridae (black fungus gnats)4. The fruitfly Drosophila is the most thoroughly studied of the Dipteran flies. The lower Dipteran fly (Nematocera), Bradysia (previously called Sciara) coprophila, diverged 200 million years ago from Drosophila, which is a "higher Dipteran" fly (Brachycera). Therefore, B. coprophila is in a favorable taxonomic position for comparative studies with D. melanogaster (Figure 1). Moreover, B. coprophila has many unique biological features that are worthy of study in their own right5,6,7. Many of these features disobey the rule of DNA constancy in which all cells of an organism have the same DNA content. In B. coprophila, (i) the paternal genome is eliminated on a monopolar spindle in male meiosis I; (ii) there is non-disjunction of the X dyad in male meiosis II; (iii) germ line-limited (L) chromosomes are eliminated from the soma; and (iv) one or two X chromosomes are eliminated in the early embryo depending on the sex of the individual. Chromosome imprinting to distinguish maternal from paternal homologs was first discovered in B. coprophila and is at play for many of these chromosome elimination events. In addition to chromosome elimination, another bypass of DNA constancy occurs via developmentally regulated, locus-specific DNA amplification at the DNA puff loci in larval salivary gland polytene chromosomes. Studies of these unique features require laboratory maintenance of B.coprophila; details of its husbandry are presented here to facilitate such studies.
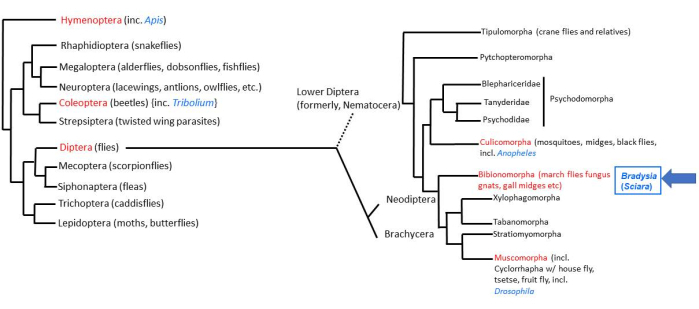
Figure 1: Phylogeny of Bradysia (Sciara) coprophila. Popular model organisms are indicated in blue font and their taxonomic order in red font. Bradysia and other Sciarid fungus gnats as well as mosquitoes are lower dipteran flies (formerly, suborder Nematocera), whereas Drosophila species are higher dipteran flies (suborder: Brachysera). Information on the left side of the figure is from Misof et al.33; information on the right side is from Bertone et al.34 and Wiegmann et al.2. Please click here to view a larger version of this figure.
Previously, the genus Sciara had the largest number (700) of species for any eukaryote, prompting Steffan to subdivide them8. Subsequently, Shin proposed that the Sciaridae family be subdivided into the subfamily Sciarinae (with six genera including Sciara, Trichosia, and Leptosciarella), the subfamiliy Megalosphyinae (including the genus Bradysia) and three other groups (including Pseudolycoriella)9. The phylogeny of the Sciaridae has been studied further by several groups in recent years9,10,11. Over the past several decades, the names of many organisms in the Sciaridae family have changed12. Although most of the literature spanning more than a century refers to the organism that we study as Sciara coprophila, its current taxonomic name is now Bradysia coprophila (syn. Bradysia tilicola and other synonyms)10. They are found worldwide and are commonly known as fungus gnats since they eat mushrooms and other fungi. They were first described in 1804 by Meigen13 in Europe and subsequently by Johannsen14,15 in North America. B. coprophila was collected at Cold Spring Harbor Laboratory and laboratory stocks were established by Charles Metz in the early 1900s when he was a graduate student at Columbia University with Thomas Hunt Morgan. Thus, the current stocks reflect a century of inbreeding. Similarly, the biology of B. coprophila was further elucidated by decades of cytogenetic studies by Helen Crouse (who did her Ph.D. work with Barbara McClintock).
In the 1930s, Bradysia (Sciara) competed with Drosophila melanogaster as a model system for genetic studies. Despite its many unique biological features, B. coprophila was eclipsed by D. melanogaster as a popular model organism since radiation-induced phenotypic mutations were needed for genetic studies and were easier to achieve in the latter, even though B. coprophila is only slightly more resistant to gamma irradiation than D. melanogaster16. In the modern era of genomics, this is no longer a concern. Since the genome sequence17,18,19 (Urban, Gerbi, and Spradling, data not shown) and methods for transformation20,21 (Yamamoto and Gerbi, data not shown) for B. coprophila have recently become available, the time is now ripe to utilize it as a new/old emerging model system, as seen by the growing community of scientists that have adopted it for their research. This article describes procedures for its laboratory maintenance.
B. coprophila lacks a Y chromosome, and the sex of the offspring is determined by the mother. Females that have the X' ("X-prime") chromosome with a long paracentric inversion will have only daughters, whereas females that are homozygous for the standard (non-inverted) X chromosome will have only sons5 (Figure 2). Sequence information is available for the X' chromosome19, but the molecular mechanism remains to be elucidated on how the X' chromosome determines that offspring will be females. Males never have the X' chromosome, and after fertilization, females are X'X (heterozygous for the X') or XX. Adult X'X females can be distinguished from XX females by phenotypic markers on the wing (Figure 3). X'X females (who will have only daughters) can be recognized by the dominant Wavy (W) wing marker on the X' (as in the HoLo2 stock)22. Alternatively, XX females (who will have only sons) can be recognized by the recessive petite (p) wing marker on the X as in the 91S stock23. In this case, X'Xp females will have full-length (not petite) wings and will have only daughters. The stock 6980 carries a recessive marker on the X chromosome for swollen (sw) veins24, as well as the dominant marker Wavy on the X', allowing two markers for selection for crosses. The degree of expression of Wavy can vary and seems weaker in overcrowded vials where food is limiting or if the temperature becomes too warm. The Wavy wing phenotype is exceptionally strong if larvae are kept in the cold room (4°-8 °C) instead of the usual 21 °C. Although the recessive petite wing marker is not variable and is very easy to identify, 91S stocks are used less frequently since they are less healthy than the HoLo2 stock. The B. coprophila mating schemes are presented here (Figure 2), and described in detail for the HoLo2, 7298, and W14 stocks (Supplemental File 1), the 91S stock (Supplemental File 1), the 6980 stock (Supplemental File 1), and the translocation stocks (Supplemental File 1). The translocation stocks are no longer extant; they were reciprocal translocations of heterochromomeres (H1, H2, and H3) on the short arm of the X that contains the ribosomal RNA genes25,26,27.
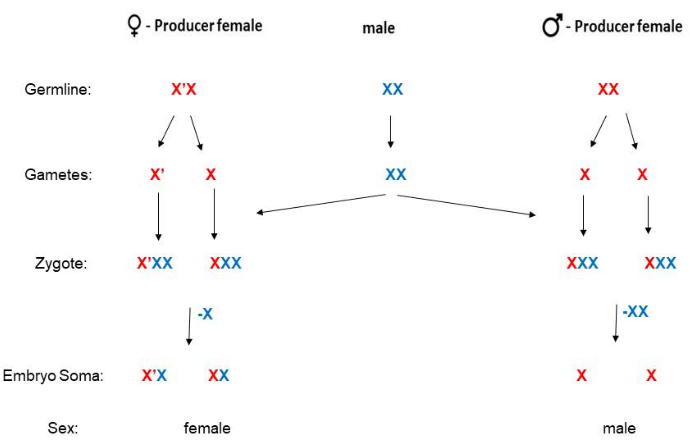
Figure 2: Mating scheme for B. coprophila. This organism has no Y chromosome (the male soma has a single X); mothers determine the sex of their offspring. XX mothers have only sons and X'X females have only daughters. The X' chromosome has a long paracentric inversion when compared to the X chromosome. The paternal or maternal lineage of the X (or X') chromosome is denoted by blue or red, respectively, in this figure. Sperm are haploid for the autosomes but have two copies of the X chromosome due to non-disjunction in meiosis II. The somatic lineage of early embryos eliminates one or two copies of the paternally derived X if they will be female or male, respectively. Please click here to view a larger version of this figure.

Figure 3: Wing phenotypes of B. coprophila. Adult female flies are shown with various wing phenotypes: (A) straight-wing (XX), (B) Wavy wing (X'WX), (C) extreme Wavy wing (X'WX) phenotype that has a shriveled appearance after storing larvae in the cold room, (D) petite wing (XpXp) that is vestigial-like, (E) straight wing with wildtype (XX) and not swollen veins, (F) straight wing with swollen veins (XswXsw) where small bubbles (black arrows) appear on the upper edge of the wing and/or near the tip of both wings, (G) extreme example of swollen where a blister (white arrow) occurs on one or both wings. Males lack the X' chromosome and therefore will never have Wavy wings, but they have petite or swollen wings in the 91S stock or 6980 stock, respectively. Scale bar = 1 mm. Please click here to view a larger version of this figure.
The goal in stock maintenance is to perform crosses where half the crosses are from female-producing mothers and half the crosses from male-producing mothers to have equal numbers of female and male adults in the next generation for subsequent crosses. However, this also involves planning since the life cycle for males is shorter than for females and adult males emerge up to a week before adult females. Nature accommodates for this asynchrony between the sexes by having the male embryos emerge as larvae 1-2 days after the female larvae from a cross on the same date. However, to ensure that male and female adults are available at the same time for laboratory crosses, female development can be somewhat speeded up by leaving vials with female larvae at room temperature rather than at 21 °C or placing vials with male larvae at slightly cooler temperatures (e.g., 16 °C). Another route that is more foolproof is to do crosses with female-producer mothers on Monday and crosses with male-producer mothers on Friday of the same week. The easiest route, which is what we employ, is to perform crosses with female-producer and male-producer mothers on the same day each week and perform crosses on that day in each consecutive week. In that approach, adult females from a cross on week 1 can be mated with adult males that emerged from a cross on week 2.
The life cycle for female B. coprophila is 5 weeks when raised at 21 °C (Table 1). The length of their life cycle is somewhat longer at cooler temperatures or if they are underfed. The life cycle of male B. coprophila is ~4-4.5 weeks since they pupate 0.5-1 week before females. The end of each larval instar is marked by the shedding of the cuticle, which is triggered by a burst in the level of the steroid hormone ecdysone. Unlike D. melanogaster, which has three larval instars, B. coprophila has four larval instars.
| Developmental Stage | Days post mating (dpm) | Length of stage (days) |
| Egg laid | 1-2 | |
| Embryo | 1-2 to 7-8 | ~7 days |
| Larva | ||
| Larval instars 1, 2, and 3 | 7-8 to 16-19 | ~10 |
| 4th larval instar pre-eyespot | 16-19 to 21-24 | 5 |
| 4th larval instar eye-spot stage | 21-24 to 25-28 | 4 |
| Pupa | 25-28 to 30-33 | 5 |
| Adult | lives 1-2 days at 21 °C if mated or lives 2-3 weeks at 16 °C if not mated. | |
Table 1: Life cycle of female B. coprophila at 21 °C.
B. coprophila can be kept anywhere in the range of 15 °C-25 °C, with development progressing more slowly at cooler temperatures. This insect prefers a humid environment (being found in the soil of houseplants or mushroom beds), so we keep a beaker with deionized water in the incubator. B. coprophila can be kept at room temperature in a metal bread box with a loosely fitting lid and containing a beaker of water, but they go into heat shock at 37 °C28, which is a danger in hot climates. Michael Ashburner and others have tried with little success to store D. melanogaster in the cold to reduce the time needed for stockkeeping. In contrast, a major advantage of B. coprophila is that vials with mid-stage larvae can be stored for up to 3 months on an open shelf in the cold room (4-8 °C) with minimal care of only feeding once a month. They develop extremely slowly in the cold up to the pupal stage and will emerge as fertile adults when the vials are brought back to 21 °C. Presumably, this mimics their overwintering in the wild. This cold-induced developmental stalling might be comparable to that seen after gamma irradiation of mid-stage B. coprophila larvae16, but developmental stalling is not seen in late-stage larvae that have passed the point of no return for their normal developmental progression.
The protocols described here have led to proven success in raising B. coprophila. When recently eclosed fat adults are chosen for mating (Figure 4), over 90% of the crosses can be fertile and yield offspring. The fertility success does vary with different stocks (Table 2). Stock 7298 (X' chromosome with Wavy marker) was the healthiest of the stocks but went through a period of decline, apparently due to activation of DNA mobile elements creating genome rearrangements32. The HoLo2 stock represents a healthy strain derived from 7298, where apparently the genome rearrangements have stabilized, and it has replaced the parental 7298 stock in the stock center. The HoLo2 stock is the one that was used to sequence the B. coprophila genome and is the one used most widely by various laboratory groups. Recently, CRISPR mutagenesis of HoLo2 flies has been used to create the W14 stock with a white eye phenotype to be used for transformation with fluorescent eye markers (Yamamoto and Gerbi, data not shown). The W14 strain is exceptionally robust. The 6980 stock (Wavy wing and swollen vein markers) is somewhat less robust and the 91S stock (petite wing marker) is even less robust.
Successful crosses result in embryos (Figure 5). Embryos undergo elimination of the imprinted paternal X chromosomes at the 7th to 9th cleavage division. In addition, germ-line limited L chromosomes are eliminated from the somatic lineage in embryos at the 5th to 6th cleavage division. The embryos emerge as larvae, which should not be confused with mold that also may be present (Figure 6). Eyespots (anlage to the adult eyes) appear at the second half of the 4th larval instar (Figure 7). The size of the eyespots provides a convenient phenotypic marker for the onset and progression of DNA puff amplification, which is one of only two known examples of naturally occurring developmentally regulated site-specific intrachromosomal DNA (gene) amplification. Subsequently, pupae develop, and the amount of pigment filling their eyes can serve as a developmental marker for meiosis I and II in spermatogenesis (Figure 8) with its unique chromosome behaviors in these divisions.

Figure 4: Mating of adult B. coprophila flies. The upper panel shows the three types of adult flies in the HoLo2 stock: female-producing mothers with Wavy wings (X'WX adult females), male-producing mothers with straight wings (XX adult females), and males with straight wings (X0 adult males). Note the pointed ovipositor at the posterior end of the female flies and the hook-shaped clasper at the posterior end of the male flies. The lower panel shows a male and female mating, where the male clasper has grabbed the female's ovipositor. The sperm will be stored in the spermatheca of the female and will fertilize the eggs as they are discharged to the outside. The length of adults is 2.0 mm (males), 2.5 mm (females). Please click here to view a larger version of this figure.
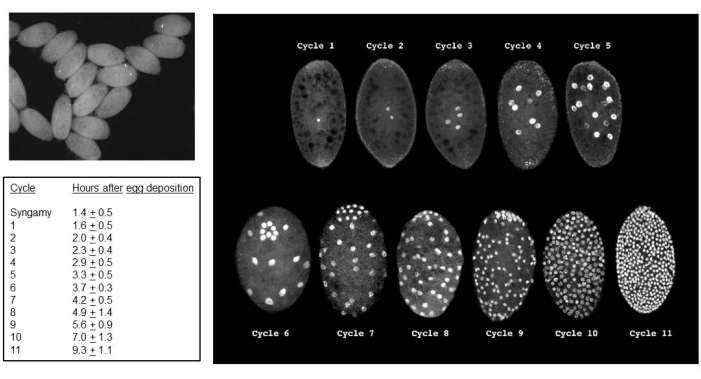
Figure 5: B. coprophila embryos. The upper left panel is a view of embryos using standard light in a dissecting microscope; the nuclei in the syncytial cytoplasm appear as black dots. The panel on the right uses fluorescence microscopy to visualize the propidium iodide-stained nuclei of embryos. The embryos have an average length of 200 microns and an average width of 150 microns. The nuclei for the germ cells cluster at the posterior pole of the embryo as seen for cycles 6 (embryo tilted forward), and 7-9, after which they are interspersed with somatic nuclei. L chromosome elimination in the somatic lineage occurs in cleavage division 5 or 6; X chromosome elimination in the somatic lineage occurs in the 7th, 8th, or 9th cleavage division. Cellularization occurs during interphase of cycle 11. The table at the left shows the average length of time for each division cycle at 22 °C. The table on the left and panel on the right are adapted with permission from de Saint Phalle and Sullivan35. Please click here to view a larger version of this figure.
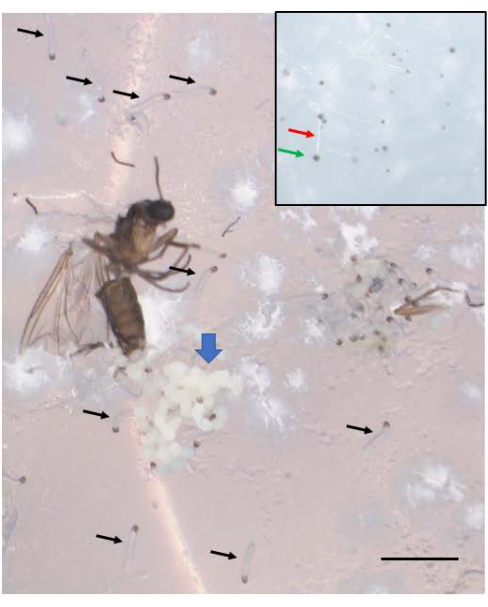
Figure 6: B. coprophila embryos emerge as larvae. A cluster of embryos (blue thick arrow) is seen near the ovipositor of the adult female who died after egg-laying. A week after egg-laying, the embryos become young larvae, several of whom are indicated by the black arrows. The newly emerged larvae have a black jaw at the anterior end and a translucent body. They move on top of the agar surface and should not be confused with mold that does not move. The inset shows some mold, with a white filament (red arrow) and a black spore at its tip (green arrow), and is very slightly smaller than the newly emerged larvae. Scale bar = 1 mm. Please click here to view a larger version of this figure.
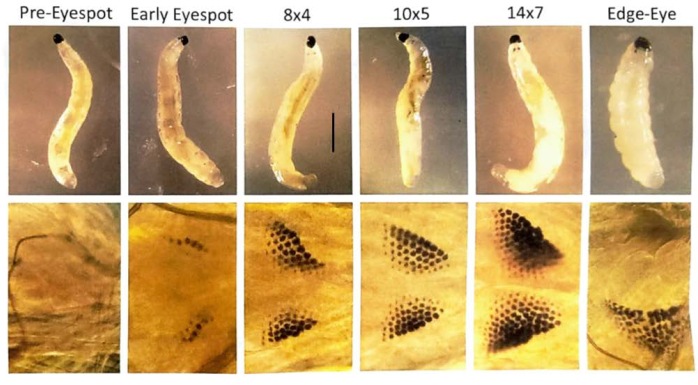
Figure 7: Eyespot stages of B. coprophila larvae. Eyespots form at the anterior of the larva, just behind the jaw, and are composed of pigment granules that increase in number. The eyespots are the anlage to the adult eye. The upper panel shows larvae that were visualized with a dissecting microscope; the lower panel is a magnified view of the eyespots using a phase contrast microscope to visualize a larva on a microscope slide with a drop of distilled water and a coverslip lightly floating on top. The nomenclature of the eyespot stages is according to Gabrusewycz-Garcia30 where the number of granules is counted in the longest row (e.g., 12) and the number of additional rows excluding the longest row is noted (e.g., 6 for eyespot stage 12×6). Initiation of site-specific DNA amplification in the salivary gland polytene chromosomes begins at eyespot stage 10×5 and is completed at 14×7 when there is a burst of transcription at the locus and expansion of the DNA puffs31. At the subsequent edge-eye/dropped jaw stage, the eyespot granules begin to merge, and they move laterally away from the midline; the length of the larval body shortens. Further, the DNA puffs condense at this stage. It takes about one day at 21 °C to traverse each eyespot stage. Scale bar = 1 mm. Please click here to view a larger version of this figure.
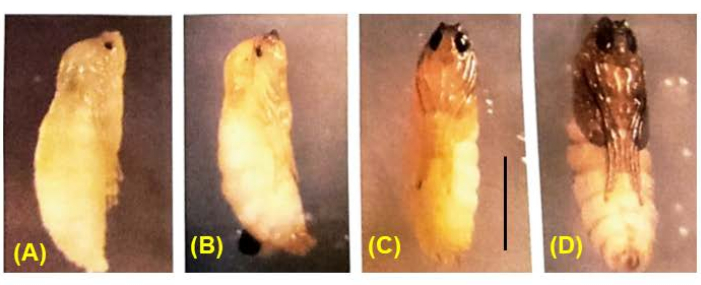
Figure 8: Development of B. coprophila pupae. During pupation, all larval tissues histolyze except for the nervous system and are replaced by adult tissues that arise by cell divisions of the imaginal discs. The body color changes from white to tan to brown to black. Pigment gradually fills in the pupal eye; meiosis I and II occur in male pupae with eyes that are 1/4 to 1/2 filled with pigment36. Scale bar = 1 mm. Please click here to view a larger version of this figure.
| Stock name | Markers | Fertility rate | Comments |
| 7298 | Wavy (W) wing | ~75% | |
| HoLo2 | Wavy (W) wing | ~90% | derived from 7298 |
| W14 | Wavy (W) wing; White eyes | ~95% | derived from HoLo2 |
| 6980 | Wavy (W) wing; swollen (sw) veins | ~65% | |
| 91S | weak Wavy (W) wing; petite (p) wings | ~50% | the Wavy marker was introduced in a cross to rescue 91S |
Table 2: Stocks of Bradysia (Sciara) coprophila. Table 1 of Gerbi6 lists these markers and others that are no longer extant. Five translocations (T1, T23, T29, T32, T70) at the centromere end of the X27 are summarized in Figure 8 of Gerbi6 but are no longer extant.
Supplemental File 1: HoLo2 (and 7298 and W14) crosses, 91S cross and rescue, 6980 cross, translocation crosses. Please click here to download this File.
