Полное понимание биологических принципов требует изучения многих разнообразных организмов, охватывающих Древо Жизни. Несмотря на то, что до конца19-го века был описан широкий спектр организмов, к середине20-го века экспериментальные исследования стали ограничены горсткой из менее чем дюжины модельных организмов. С наступлением эры генома и целью секвенирования геномов всех видов в Древе Жизни1 мы теперь в состоянии расширить типы организмов, используемых для лабораторных экспериментов, и извлечь выгоду из их разнообразия. Такое расширение новых модельных организмов для экспериментов имеет предпосылку для возможности поддерживать их в лаборатории. Здесь описываются протоколы для выращивания одного из таких появляющихся новых/старых модельных организмов.
Большая часть животной жизни на Земле приходится на четыре сверхизлучения насекомых2. Среди насекомых насчитывается около 158 000 видов двукрылых (настоящих мух)3, из которых около 3000 видов входят в семейство Sciaridae (черные грибные комары)4. Плодовая муха дрозофила является наиболее тщательно изученной из двукрылых мух. Низшая двукрылая муха (Nematocera), Bradysia (ранее называвшаяся Sciara) coprophila, 200 миллионов лет назад отделилась от дрозофилы, которая является «высшей двукрылой» мухой (Brachycera). Таким образом, B. coprophila находится в выгодном таксономическом положении для сравнительных исследований с D. melanogaster (рис. 1). Кроме того, B. coprophila обладает многими уникальными биологическими особенностями, которые сами по себе заслуживают изучения 5,6,7. Многие из этих особенностей нарушают правило постоянства ДНК, согласно которому все клетки организма имеют одинаковое содержание ДНК. У B. coprophila (i) отцовский геном элиминируется на монополярном веретене при мужском мейозе I; (ii) при мужском мейозе II наблюдается недизъюнкция диады X; (iii) хромосомы, ограниченные зародышевой линией (L), элиминируются из сомы; и (iv) одна или две Х-хромосомы элиминируются на ранних стадиях развития эмбриона в зависимости от пола особи. Импринтинг хромосом для различения материнских и отцовских гомологов был впервые обнаружен у B. coprophila и играет роль во многих из этих событий элиминации хромосом. В дополнение к элиминации хромосом, еще один обход постоянства ДНК происходит посредством регулируемой в развитии, локус-специфичной амплификации ДНК в локусах затяжки ДНК в политенных хромосомах личиночных слюнных желез. Изучение этих уникальных особенностей требует лабораторного сопровождения B.coprophila; Подробности его содержания представлены здесь для облегчения таких исследований.
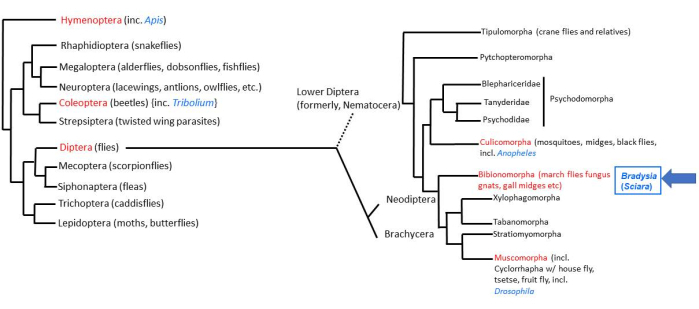
Рисунок 1: Филогения Bradysia (Sciara) coprophila. Популярные модельные организмы обозначены синим шрифтом, а их таксономический порядок — красным. Bradysia и другие комары Sciarid, а также комары относятся к низшим двукрылым (ранее подотряд Nematocera), тогда как виды дрозофил относятся к высшим двукрылым (подотряд: Brachysera). Информация в левой части рисунка взята из Misof et al.33; информация справа взята из Bertone et al.34 и Wiegmann et al.2. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
Ранее род Sciara имел самое большое количество (700) видов среди всех эукариот, что побудило Стеффана разделить ихна 8. Впоследствии Шин предложил разделить семейство Sciaridae на подсемейство Sciarinae (с шестью родами, включающими Sciara, Trichosia и Leptosciarella), семейство Megalosphyinae (включая род Bradysia) и три другие группы (включая Pseudolycoriella)9. Филогения Sciaridae была дополнительно изучена несколькими группами в последние годы 9,10,11. За последние несколько десятилетий названия многих организмов семейства Sciaridae изменились12. Хотя большая часть литературы, охватывающей более ста лет, называет организм, который мы изучаем, Sciara coprophila, его нынешнее таксономическое название теперь Bradysia coprophila (син. Bradysia tilicola и другие синонимы)10. Они встречаются по всему миру и широко известны как грибные комары, поскольку едят грибы и другие грибы. Впервые они были описаны в 1804 году Мейгеном13 в Европе, а затем Иогансеном 14,15 в Северной Америке. B. coprophila была собрана в лаборатории Колд-Спринг-Харбор, а лабораторные запасы были созданы Чарльзом Метцем в начале 1900-х годов, когда он был аспирантом Колумбийского университета вместе с Томасом Хантом Морганом. Таким образом, нынешние поголовья отражают столетие инбридинга. Аналогичным образом, биология B. coprophila была дополнительно прояснена в результате десятилетий цитогенетических исследований Хелен Крауз (которая работала над докторской диссертацией вместе с Барбарой Мак-Клинток).
В 1930-х годах Bradysia (Sciara) конкурировала с Drosophila melanogaster в качестве модельной системы для генетических исследований. Несмотря на множество уникальных биологических особенностей, B. coprophila затмила D. melanogaster в качестве популярного модельного организма, поскольку радиационно-индуцированные фенотипические мутации были необходимы для генетических исследований и их было легче достичь в последнем, даже несмотря на то, что B. coprophila лишь немного более устойчива к гамма-облучению, чем D. melanogaster16. В современную эпоху геномики это уже не вызывает беспокойства. С тех пор как последовательность генома 17,18,19 (Урбан, Герби и Спрэдлинг, данные не показаны) и методы трансформации 20,21 (Ямамото и Герби, данные не показаны) для B. coprophila стали недавно, настало время использовать ее в качестве новой/старой модельной системы, как это видит растущее сообщество ученых, которые приняли ее в своих исследованиях. В данной статье описаны процедуры его лабораторного обслуживания.
У B. coprophila отсутствует Y-хромосома, а пол потомства определяется по матери. У самок с X’ («Х-прайм») хромосомой с длинной парацентрической инверсией будут только дочери, тогда как у самок, гомозиготных по стандартной (неинвертированной) Х-хромосоме, будет только 5 сыновей(рис. 2). Информация о последовательностях доступна для19-й Х-хромосомы, но молекулярный механизм еще предстоит выяснить, как Х-хромосома определяет, что потомство будет самкой. Самцы никогда не имеют X’-хромосомы, а после оплодотворения самки становятся X’X (гетерозиготными по X’) или XX. Взрослых самок X’X можно отличить от самок XX по фенотипическим маркерам на крыле (рис. 3). Самки X’X (у которых будут только дочери) могут быть распознаны по доминирующему маркеру волнистого (W) крыла на X’ (как у популяции HoLo2)22. В качестве альтернативы, самки XX (у которых будут только сыновья) могут быть распознаны по рецессивному миниатюрному (p) маркеру крыла на X, как в 91S stock23. В этом случае самки X’Xp будут иметь полноразмерные (не миниатюрные) крылья и будут иметь только дочерей. Сток 6980 несет рецессивный маркер на Х-хромосоме для опухших (sw) вен24, а также доминантный маркер Wavy на X’, позволяющий использовать два маркера для отбора для скрещивания. Степень экспрессии волнистого может варьироваться и кажется слабее в переполненных флаконах, где количество пищи ограничено или если температура становится слишком высокой. Фенотип волнистого крыла особенно силен, если личинки содержатся в холодном помещении (4°-8 °C) вместо обычных 21 °C. Несмотря на то, что рецессивный маркер малого крыла не является изменчивым и его очень легко идентифицировать, стаи 91S используются реже, поскольку они менее здоровы, чем стадо HoLo2. Схемы спаривания B. coprophila представлены здесь (Рисунок 2) и подробно описаны для популяций HoLo2, 7298 и W14 (Дополнительный файл 1), 91S (Дополнительная папка 1), 6980 (Дополнительная папка 1) и транслокационных популяций (Дополнительный файл 1). Запасы транслокаций больше не сохранились; они представляли собой реципрокные транслокации гетерохромомеров (H1, H2 и H3) на коротком плече X-хромомера, содержащем гены рибосомной РНК 25,26,27.
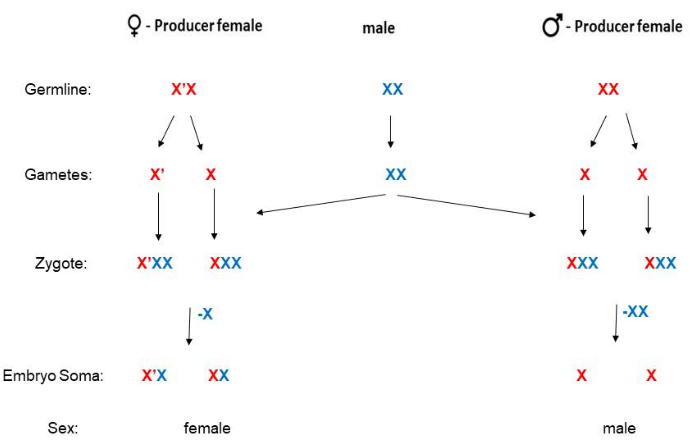
Рисунок 2: Схема спаривания B. coprophila. У этого организма нет Y-хромосомы (у самца сомы есть одна X); Матери определяют пол своего потомства. У матерей XX только сыновья, а у женщин X’X только дочери. Х-хромосома имеет длинную парацентрическую инверсию по сравнению с Х-хромосомой. Отцовская или материнская линия Х-хромосомы (или Х’) на этом рисунке обозначена синим или красным цветом соответственно. Сперматозоиды гаплоидны для аутосом, но имеют две копии Х-хромосомы из-за нерасхождения при мейозе II. Соматическая линия ранних эмбрионов исключает одну или две копии Х-хромосом, полученных по отцовской линии, если они будут женского или мужского пола соответственно. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.

Рисунок 3: Фенотипы крыльев B. coprophila. Взрослые самки мух показаны с различными фенотипами крыльев: (А) прямокрылые (XX), (В) волнистые (X’WX), (В) экстремальные волнистые крылья (X’WX) фенотипы, которые имеют сморщенный вид после накопления личинок в холодном room, (D) миниатюрные крылы (XpXp), которые являются рудиментарными, (E) прямое крыло с диким типом (XX) и не вздутыми жилками, (F) прямое крыло с вздутыми прожилками (XswXsw), где маленькие пузырьки (черные стрелки) появляются на верхнем крае крыла и/или возле кончика обоих крыльев, (G) крайний пример вздутия, когда волдырь (белая стрелка) встречается на одном или обоих крыльях. У самцов отсутствует X-хромосома, и поэтому у них никогда не будет волнистых крыльев, но у них есть миниатюрные или опухшие крылья у 91S или 6980 соответственно. Масштабная линейка = 1 мм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
Цель содержания поголовья состоит в том, чтобы проводить скрещивания, в которых половина скрещиваний происходит от матерей-самок, а половина скрещивания от матерей-производителей-самцов, чтобы иметь равное количество взрослых самок и самцов в следующем поколении для последующих скрещиваний. Однако это также включает в себя планирование, поскольку жизненный цикл у самцов короче, чем у самок, а взрослые самцы появляются на неделю раньше взрослых самок. Природа приспосабливается к этой асинхронности между полами, заставляя мужские эмбрионы появляться в виде личинок через 1-2 дня после женских личинок от скрещивания в ту же дату. Тем не менее, чтобы гарантировать, что взрослые особи самцов и самки доступны одновременно для лабораторных скрещиваний, развитие самок может быть несколько ускорено, если оставлять флаконы с личинками самок при комнатной температуре, а не при 21 °C, или размещать флаконы с личинками самцов при несколько более низких температурах (например, 16 °C). Другой способ, который является более надежным, – это скрещивание с матерями-женщинами-производителями в понедельник и скрещивание с матерями-мужчинами-производителями в пятницу на той же неделе. Самый простой путь, который мы используем, заключается в том, чтобы проводить скрещивание с самками-производителями и самцами-производителями в один и тот же день каждую неделю и выполнять скрещивание в этот день в течение каждой последующей недели. При таком подходе взрослые самки от скрещивания на 1-й неделе могут быть спарены со взрослыми самцами, которые появились от скрещивания на 2-й неделе.
Жизненный цикл самки B. coprophila составляет 5 недель при выращивании при 21 °C (Таблица 1). Продолжительность их жизненного цикла несколько увеличивается при более низких температурах или при недостаточном кормлении. Жизненный цикл самцов B. coprophila составляет ~4-4,5 недели, так как они окукливаются на 0,5-1 неделю раньше самок. Окончание каждой личиночной стадии отмечается сбрасыванием кутикулы, которое провоцируется всплеском уровня стероидного гормона экдизона. В отличие от D. melanogaster, у которого три личиночных возраста, у B. coprophila четыре личиночных возраста.
| Стадия развития | Количество дней после вязки (dpm) | Продолжительность этапа (дней) |
| Снесенное яйцо | 1-2 | |
| Эмбрион | С 1-2 по 7-8 | ~7 дней |
| Личинка | ||
| Возрасты личинок 1, 2 и 3 | с 7-8 по 16-19 | ~10 |
| Предглазное пятно 4-го личиночного возраста | с 16-19 по 21-24 | 5 |
| 4-я стадия пятен глазного пятна личиночного возраста | с 21-24 по 25-28 | 4 |
| Куколка | с 25-28 по 30-33 | 5 |
| Взрослый | живет 1-2 дня при 21 °C при спаривании или 2-3 недели при 16 °C при отсутствии спаривания. | |
Таблица 1: Жизненный цикл самок B. coprophila при 21 °C.
B. coprophila может содержаться в любом месте в диапазоне от 15 °C до 25 °C, при этом развитие протекает медленнее при более низких температурах. Это насекомое предпочитает влажную среду (находящееся в почве комнатных растений или грибных грядок), поэтому мы держим в инкубаторе стакан с деионизированной водой. B. coprophila можно хранить при комнатной температуре в металлической хлебнице со слабо прилегающей крышкой, содержащей стакан с водой, но они подвергаются тепловому шоку при 37 °C28, что представляет опасность в жарком климате. Майкл Эшбернер и другие безуспешно пытались хранить D. melanogaster в холоде, чтобы сократить время, необходимое для хранения запасов. В отличие от этого, основным преимуществом B. coprophila является то, что флаконы с личинками средней стадии могут храниться до 3 месяцев на открытой полке в холодном помещении (4-8 °C) с минимальным уходом, кормлением только один раз в месяц. Они развиваются чрезвычайно медленно на холоде вплоть до стадии куколки и становятся фертильными взрослыми особями, когда флаконы возвращаются к температуре 21 °C. Предположительно, это имитирует их зимовку в дикой природе. Эта задержка развития, вызванная холодом, может быть сравнима с той, которая наблюдается после гамма-облучения личинок B. coprophila 16 на средней стадии, но задержка развития не наблюдается у личинок поздних стадий, которые прошли точку невозврата для их нормального прогресса в развитии.
