A complete understanding of biological principles requires the study of many diverse organisms spanning the Tree of Life. Although a broad range of organisms were described through the end of the 19th century, by the middle of the 20th century experimental studies became restricted to a handful of less than a dozen model organisms. With the advent of the genomic era and the goal to sequence genomes from all species in the Tree of Life1, we are now in a position to expand the types of organisms being used for laboratory experiments and to reap the advantage of their diversity. Such expansion of emerging model organisms for experiments has a prerequisite of being able to maintain them in the laboratory. Here, protocols are described for rearing one such emerging new/old model organism.
The majority of animal life on Earth is accounted for by four super-radiations of Insects2. Within the Insects, there are about 158,000 species of Diptera (true flies)3, with about 3000 species in the family Sciaridae (black fungus gnats)4. The fruitfly Drosophila is the most thoroughly studied of the Dipteran flies. The lower Dipteran fly (Nematocera), Bradysia (previously called Sciara) coprophila, diverged 200 million years ago from Drosophila, which is a "higher Dipteran" fly (Brachycera). Therefore, B. coprophila is in a favorable taxonomic position for comparative studies with D. melanogaster (Figure 1). Moreover, B. coprophila has many unique biological features that are worthy of study in their own right5,6,7. Many of these features disobey the rule of DNA constancy in which all cells of an organism have the same DNA content. In B. coprophila, (i) the paternal genome is eliminated on a monopolar spindle in male meiosis I; (ii) there is non-disjunction of the X dyad in male meiosis II; (iii) germ line-limited (L) chromosomes are eliminated from the soma; and (iv) one or two X chromosomes are eliminated in the early embryo depending on the sex of the individual. Chromosome imprinting to distinguish maternal from paternal homologs was first discovered in B. coprophila and is at play for many of these chromosome elimination events. In addition to chromosome elimination, another bypass of DNA constancy occurs via developmentally regulated, locus-specific DNA amplification at the DNA puff loci in larval salivary gland polytene chromosomes. Studies of these unique features require laboratory maintenance of B.coprophila; details of its husbandry are presented here to facilitate such studies.
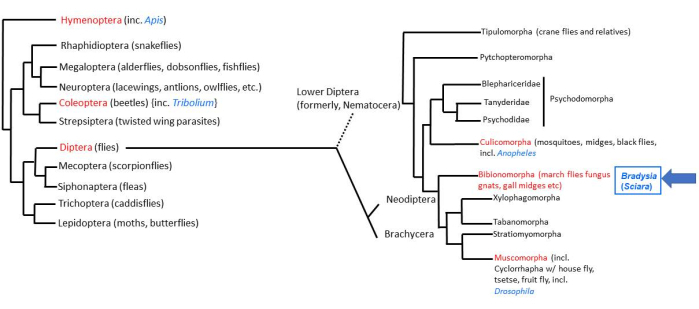
Figure 1: Phylogeny of Bradysia (Sciara) coprophila. Popular model organisms are indicated in blue font and their taxonomic order in red font. Bradysia and other Sciarid fungus gnats as well as mosquitoes are lower dipteran flies (formerly, suborder Nematocera), whereas Drosophila species are higher dipteran flies (suborder: Brachysera). Information on the left side of the figure is from Misof et al.33; information on the right side is from Bertone et al.34 and Wiegmann et al.2. Please click here to view a larger version of this figure.
Previously, the genus Sciara had the largest number (700) of species for any eukaryote, prompting Steffan to subdivide them8. Subsequently, Shin proposed that the Sciaridae family be subdivided into the subfamily Sciarinae (with six genera including Sciara, Trichosia, and Leptosciarella), the subfamiliy Megalosphyinae (including the genus Bradysia) and three other groups (including Pseudolycoriella)9. The phylogeny of the Sciaridae has been studied further by several groups in recent years9,10,11. Over the past several decades, the names of many organisms in the Sciaridae family have changed12. Although most of the literature spanning more than a century refers to the organism that we study as Sciara coprophila, its current taxonomic name is now Bradysia coprophila (syn. Bradysia tilicola and other synonyms)10. They are found worldwide and are commonly known as fungus gnats since they eat mushrooms and other fungi. They were first described in 1804 by Meigen13 in Europe and subsequently by Johannsen14,15 in North America. B. coprophila was collected at Cold Spring Harbor Laboratory and laboratory stocks were established by Charles Metz in the early 1900s when he was a graduate student at Columbia University with Thomas Hunt Morgan. Thus, the current stocks reflect a century of inbreeding. Similarly, the biology of B. coprophila was further elucidated by decades of cytogenetic studies by Helen Crouse (who did her Ph.D. work with Barbara McClintock).
In the 1930s, Bradysia (Sciara) competed with Drosophila melanogaster as a model system for genetic studies. Despite its many unique biological features, B. coprophila was eclipsed by D. melanogaster as a popular model organism since radiation-induced phenotypic mutations were needed for genetic studies and were easier to achieve in the latter, even though B. coprophila is only slightly more resistant to gamma irradiation than D. melanogaster16. In the modern era of genomics, this is no longer a concern. Since the genome sequence17,18,19 (Urban, Gerbi, and Spradling, data not shown) and methods for transformation20,21 (Yamamoto and Gerbi, data not shown) for B. coprophila have recently become available, the time is now ripe to utilize it as a new/old emerging model system, as seen by the growing community of scientists that have adopted it for their research. This article describes procedures for its laboratory maintenance.
B. coprophila lacks a Y chromosome, and the sex of the offspring is determined by the mother. Females that have the X' ("X-prime") chromosome with a long paracentric inversion will have only daughters, whereas females that are homozygous for the standard (non-inverted) X chromosome will have only sons5 (Figure 2). Sequence information is available for the X' chromosome19, but the molecular mechanism remains to be elucidated on how the X' chromosome determines that offspring will be females. Males never have the X' chromosome, and after fertilization, females are X'X (heterozygous for the X') or XX. Adult X'X females can be distinguished from XX females by phenotypic markers on the wing (Figure 3). X'X females (who will have only daughters) can be recognized by the dominant Wavy (W) wing marker on the X' (as in the HoLo2 stock)22. Alternatively, XX females (who will have only sons) can be recognized by the recessive petite (p) wing marker on the X as in the 91S stock23. In this case, X'Xp females will have full-length (not petite) wings and will have only daughters. The stock 6980 carries a recessive marker on the X chromosome for swollen (sw) veins24, as well as the dominant marker Wavy on the X', allowing two markers for selection for crosses. The degree of expression of Wavy can vary and seems weaker in overcrowded vials where food is limiting or if the temperature becomes too warm. The Wavy wing phenotype is exceptionally strong if larvae are kept in the cold room (4°-8 °C) instead of the usual 21 °C. Although the recessive petite wing marker is not variable and is very easy to identify, 91S stocks are used less frequently since they are less healthy than the HoLo2 stock. The B. coprophila mating schemes are presented here (Figure 2), and described in detail for the HoLo2, 7298, and W14 stocks (Supplemental File 1), the 91S stock (Supplemental File 1), the 6980 stock (Supplemental File 1), and the translocation stocks (Supplemental File 1). The translocation stocks are no longer extant; they were reciprocal translocations of heterochromomeres (H1, H2, and H3) on the short arm of the X that contains the ribosomal RNA genes25,26,27.
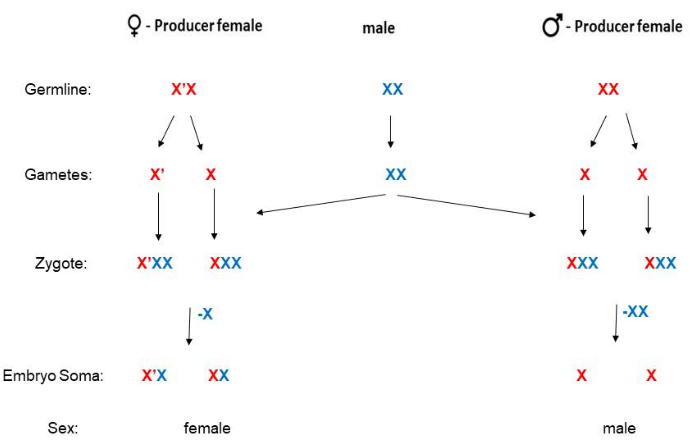
Figure 2: Mating scheme for B. coprophila. This organism has no Y chromosome (the male soma has a single X); mothers determine the sex of their offspring. XX mothers have only sons and X'X females have only daughters. The X' chromosome has a long paracentric inversion when compared to the X chromosome. The paternal or maternal lineage of the X (or X') chromosome is denoted by blue or red, respectively, in this figure. Sperm are haploid for the autosomes but have two copies of the X chromosome due to non-disjunction in meiosis II. The somatic lineage of early embryos eliminates one or two copies of the paternally derived X if they will be female or male, respectively. Please click here to view a larger version of this figure.

Figure 3: Wing phenotypes of B. coprophila. Adult female flies are shown with various wing phenotypes: (A) straight-wing (XX), (B) Wavy wing (X'WX), (C) extreme Wavy wing (X'WX) phenotype that has a shriveled appearance after storing larvae in the cold room, (D) petite wing (XpXp) that is vestigial-like, (E) straight wing with wildtype (XX) and not swollen veins, (F) straight wing with swollen veins (XswXsw) where small bubbles (black arrows) appear on the upper edge of the wing and/or near the tip of both wings, (G) extreme example of swollen where a blister (white arrow) occurs on one or both wings. Males lack the X' chromosome and therefore will never have Wavy wings, but they have petite or swollen wings in the 91S stock or 6980 stock, respectively. Scale bar = 1 mm. Please click here to view a larger version of this figure.
The goal in stock maintenance is to perform crosses where half the crosses are from female-producing mothers and half the crosses from male-producing mothers to have equal numbers of female and male adults in the next generation for subsequent crosses. However, this also involves planning since the life cycle for males is shorter than for females and adult males emerge up to a week before adult females. Nature accommodates for this asynchrony between the sexes by having the male embryos emerge as larvae 1-2 days after the female larvae from a cross on the same date. However, to ensure that male and female adults are available at the same time for laboratory crosses, female development can be somewhat speeded up by leaving vials with female larvae at room temperature rather than at 21 °C or placing vials with male larvae at slightly cooler temperatures (e.g., 16 °C). Another route that is more foolproof is to do crosses with female-producer mothers on Monday and crosses with male-producer mothers on Friday of the same week. The easiest route, which is what we employ, is to perform crosses with female-producer and male-producer mothers on the same day each week and perform crosses on that day in each consecutive week. In that approach, adult females from a cross on week 1 can be mated with adult males that emerged from a cross on week 2.
The life cycle for female B. coprophila is 5 weeks when raised at 21 °C (Table 1). The length of their life cycle is somewhat longer at cooler temperatures or if they are underfed. The life cycle of male B. coprophila is ~4-4.5 weeks since they pupate 0.5-1 week before females. The end of each larval instar is marked by the shedding of the cuticle, which is triggered by a burst in the level of the steroid hormone ecdysone. Unlike D. melanogaster, which has three larval instars, B. coprophila has four larval instars.
| Developmental Stage | Days post mating (dpm) | Length of stage (days) |
| Egg laid | 1-2 | |
| Embryo | 1-2 to 7-8 | ~7 days |
| Larva | ||
| Larval instars 1, 2, and 3 | 7-8 to 16-19 | ~10 |
| 4th larval instar pre-eyespot | 16-19 to 21-24 | 5 |
| 4th larval instar eye-spot stage | 21-24 to 25-28 | 4 |
| Pupa | 25-28 to 30-33 | 5 |
| Adult | lives 1-2 days at 21 °C if mated or lives 2-3 weeks at 16 °C if not mated. | |
Table 1: Life cycle of female B. coprophila at 21 °C.
B. coprophila can be kept anywhere in the range of 15 °C-25 °C, with development progressing more slowly at cooler temperatures. This insect prefers a humid environment (being found in the soil of houseplants or mushroom beds), so we keep a beaker with deionized water in the incubator. B. coprophila can be kept at room temperature in a metal bread box with a loosely fitting lid and containing a beaker of water, but they go into heat shock at 37 °C28, which is a danger in hot climates. Michael Ashburner and others have tried with little success to store D. melanogaster in the cold to reduce the time needed for stockkeeping. In contrast, a major advantage of B. coprophila is that vials with mid-stage larvae can be stored for up to 3 months on an open shelf in the cold room (4-8 °C) with minimal care of only feeding once a month. They develop extremely slowly in the cold up to the pupal stage and will emerge as fertile adults when the vials are brought back to 21 °C. Presumably, this mimics their overwintering in the wild. This cold-induced developmental stalling might be comparable to that seen after gamma irradiation of mid-stage B. coprophila larvae16, but developmental stalling is not seen in late-stage larvae that have passed the point of no return for their normal developmental progression.
