Анализ роста зависит от набора инструментов, которые обычно используются учеными для описания растений генотип определяют различия роста и / или ответы фенотипические к факторам окружающей среды. Они включают в себя измерения размера и веса всего растения или органа и расчеты темпов роста, чтобы изучить основные механизмы роста. роста органов определяется делением клеток и расширения на клеточном уровне. Поэтому, в том числе количественной оценки этих двух процессов роста анализов является ключом к пониманию различий в росте целого органа 1. Следовательно, крайне важно иметь соответствующую методологию для определения клеточных параметров роста, что является относительно простым в использовании, не специализированных лабораториях.
Кинематический анализ уже создан в качестве подхода , обеспечивающего мощную основу для разработки моделей роста органа 2. Методика была оптимизирована для линейных систем,такие как Резуховидка Таля корней и листьев однодольных, но и для нелинейных систем, таких как двудольных листьев 3. В настоящее время эта методика чаще используется для изучения того, как генетические, гормональные, развития, а также факторы окружающей среды влияют на деление клеток и расширение в различных органах (Таблица 1). Кроме того, он также обеспечивает основу для связи клеточных процессов их основных биохимических, молекулярных и физиологических норм (таблица 2), хотя ограничения могут быть наложены по размеру органа и пространственной организации для методов , которые требуют больших количеств растительного материала (например, метаболит измерения, протеомики и т.д.).
Однодольные листья, такие как кукуруза (Zea Мейс – ) листа, представляют собой линейные системы , в которых клетки перемещаются от основания листа к кончику, последовательно проходя через меристемы и удлинение зоны , чтобы достичь зрелогозона. Это делает его идеальной моделью системы для количественных исследований пространственных закономерностей роста 4. Кроме того, листья кукурузы имеют большие зоны роста (меристемы и относительное удлинение зоны , занимающие несколько сантиметров 5) и предоставляют возможности для проведения исследований на других организационных уровнях. Это позволяет при исследовании (мнимых) регуляторных механизмов , контролирующих деление клеток и расширения, количественно с помощью кинематического анализа с помощью ряда молекулярных методов, физиологических измерений и клеточной биологии подходов (таблица 2).
Здесь мы приводим протокол для выполнения анализа кинематики в однодольных листьях. Во-первых, мы объясним, как провести надлежащее анализ как клеточного деления и удлинения клеток в зависимости от положения вдоль оси листа и, как вычислить кинематических параметров. Во-вторых, мы также показывают, как это может быть использовано в качестве основы для формирования выборки. Здесь мы рассмотрим два случая: в высоком разрешении по отбору пробd сосредоточены выборки, позволяя улучшить интерпретацию данных и экономии времени / денег, соответственно.
Таблица 1. Обзор кинематических методы анализа для количественного определения клеточного деления и расширения в различных органах.
| орган | Справка |
| однодольные листья | 16, 20, 21, 22 |
| кончики корней | 2, 23, 24, 25, 26, 27, 28, 29 |
| двудольные листья | 21, 30, 31 |
| стрелять апикальной меристемы | 32 |
Таблица 1. Обзор кинематических методы анализа для количественного определения клеточного деления и расширения в различных органах.
<p class="jove_content" fo:keep-together.within-страница = "1">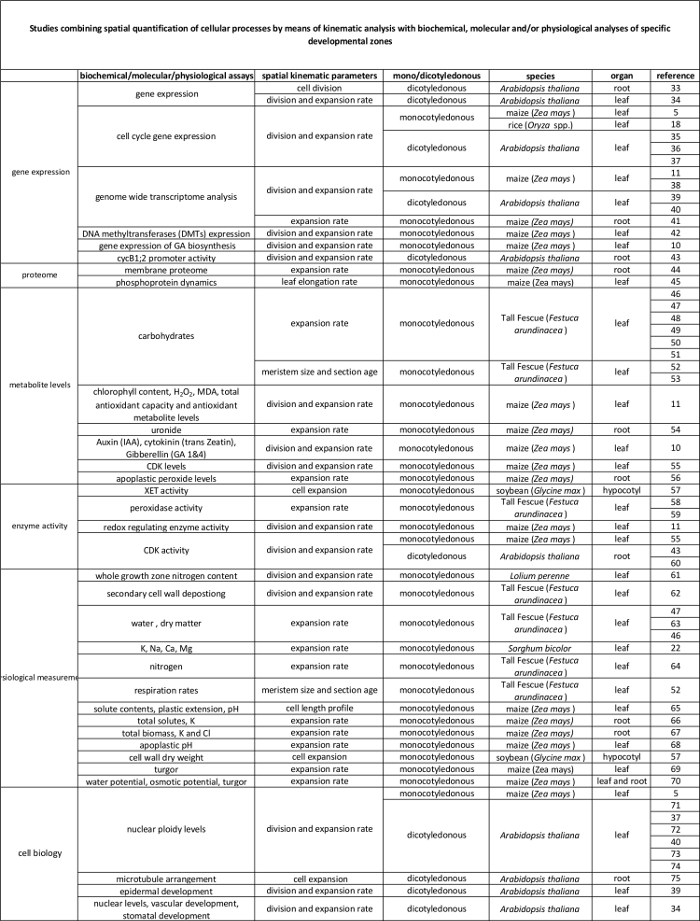
Таблица 2. Связь между клеточными процессами количественно с помощью кинематического анализа их регуляции на молекулярном уровне. Ссылки на различных исследований , связывающих количественное клеточных процессов на результатах биохимических и молекулярных анализов в различных видах и органах. Ксилоглюкан endotransglucosylase (XET), малонового (MDA), циклин-зависимых киназ (CDK). Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой таблицы.
