Serum DHT levels and Glucose tolerance test
DHT levels were measured from collected serum by both ELISA and by LC-MS according to protocol 1.24–1.25, and 2.9, 3.0. The DHT absolute values are different between mass spectrometry and ELISA, however, the relative fold (around 2-fold) of DHT vs no-DHT insertion is similar from both assays and across experiments15,23,24 (Figure 2A). DHT levels are significantly increased from preconception through pregnancy in both DHT and no-DHT mice, however, DHT levels are 2-fold higher in the DHT mice compared to the no-DHT mice at both pregestational (one day before mating) and gestational (around 14 days) time points (Figure 2B). Since absolute levels of DHT differ between ELISA and LC-MS, we calculate relative levels (fold change: DHT level with DHT insertion vs no-DHT insertion) within assays. Female mice with DHT showed impaired glucose tolerance relative to no-DHT (Figure 3) at 2 weeks following DHT insertion according to protocol 1.23.
Female offspring body weight and puberty
While generally infertile, some dams get pregnant and have pups (around 30% pregnancy rate, so testing fertility of 10 dams we will usually get 3 pregnant dams, refer to protocol 2.1). We therefore can evaluate chronic androgen effects in dams on the female offspring. Female offspring from DHT inserted dams are called DHT-daughters, and those from no-DHT inserted dams are called no-DHT daughters. We observed no difference between no-DHT and DHT-daughters for the age of vaginal opening. However, the first estrus of DHT-daughters is significantly delayed (day 35 for no-DHT daughter; day 42 for DHT-daughter). This is associated with reduced body weight at both 35 and 42 PND in DHT-daughters relative to no-DHT daughters (Figure 4; refer to protocol 2.4–2.7).
Hormonal levels and estrous cyclicity in female offspring
Testosterone levels are not altered at 21 PND, but are reduced at 26PND. However, testosterone is increased at 70 PND (Figure 5, refer to protocol 2.9–3.0). Adult DHT daughters showed disrupted estrous cyclicity compared to adult no-DHT daughters, experiencing significantly longer times in M/D and less time in P and E (Figure 6A, B; according to the method in protocol 1.17–1.22).
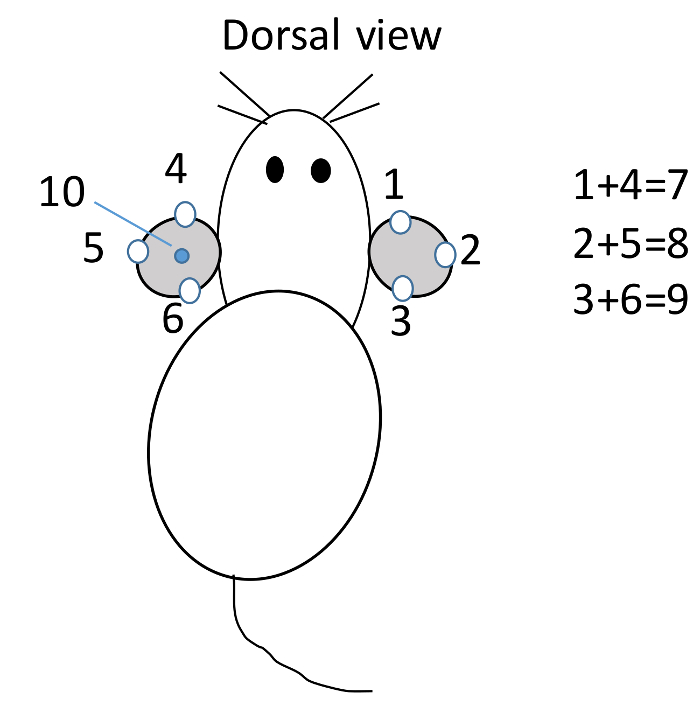
Figure 1: Mouse identification. Within one cage, mouse is ear punched at different position to represent the mouse number. Mouse #1 to 3 is punched on the right ear, and #4 to 6 is punched on the left ear viewed dorsally. If mouse is punched on ears at both #1 and #4 position, it is #7; at 2 and 5, is #8; at 3 and 6 is #9; at the middle of left ear is #10; at 1 and 10 position is #11; etc. Please click here to view a larger version of this figure.
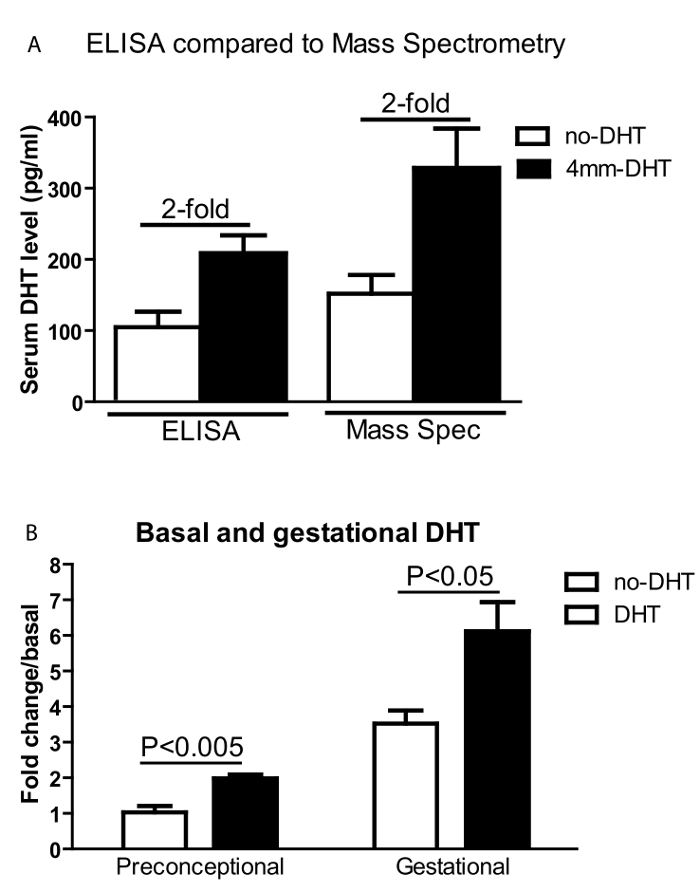
Figure 2: Serum DHT levels. Serum DHT levels. (A) Serum DHT levels are measured by both ELISA and Mass-Spectrometry. Although the absolute values are different, the two assays showed similar fold differences between DHT and no-DHT treated mice. (B) Serum DHT fold change relative to non-DHT levels at preconceptional and gestational time points. No-DHT (open bars) and DHT-implanted (black bars) female mice before (one day before mating) and during pregnancy (around 14 days of gestation). Values are mean ±S.E.M. N = 5-8 per group. Please click here to view a larger version of this figure.
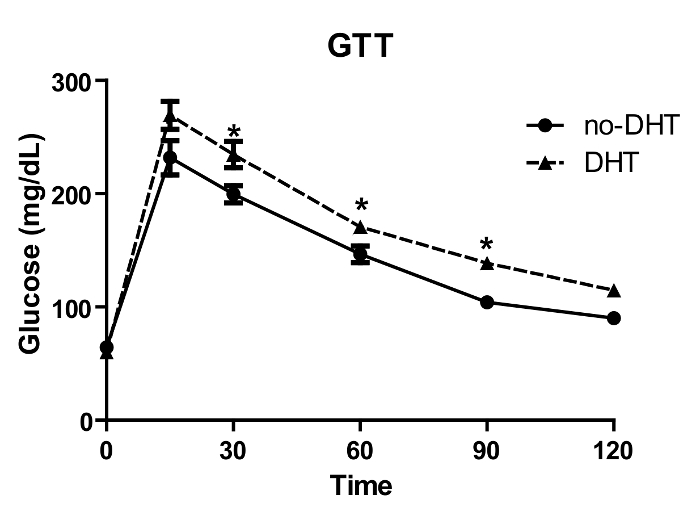
Figure 3: Glucose tolerance test (GTT). No-DHT and DHT implanted mice were fasted overnight and glucose (2 g/kg BW) was injected intraperitoneally, and tail blood glucose was measured at different time points. DHT treated mice showed significantly increased glucose levels between 30 to 120 min compared to no-DHT treated mice. Values are mean ±S.E.M. N = 4-12 per group. * P <0.05 Please click here to view a larger version of this figure.
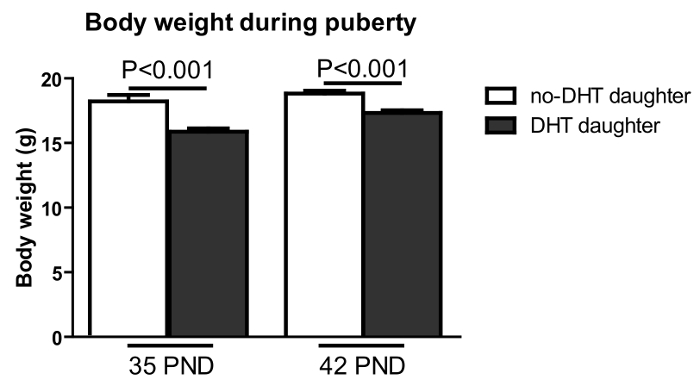
Figure 4: Pre-pregnancy maternal DHT treatment resulted in reduced body weight in female offspring at 35 and 42 PND. Body weight (Y-axis) was measured on post-natal days as shown (X-axis). DHT-exposed offspring: black bar; no –DHT offspring: open bar. Values are mean ±S.E.M. N = 9-14 per group. Please click here to view a larger version of this figure.
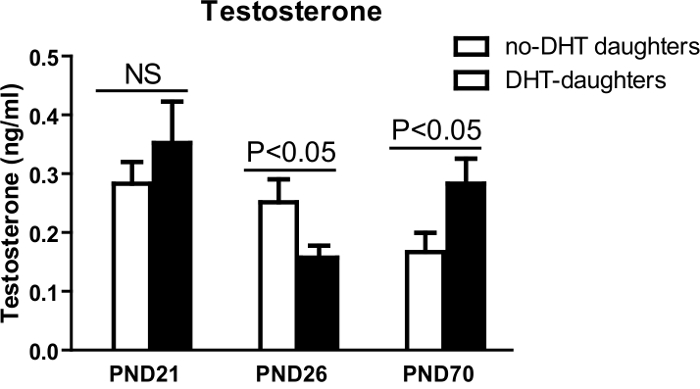
Figure 5: Serum DHT levels. Blood was collected in the morning on PND 21, 26, 70 in the morning. Serum DHT levels (Y-axis) from female offspring of no-DHT daughters (open bars) and DHT-daughters (black bars). Values are mean ±S.E.M. N = 5-11 per group. This figure has been modified from reference2 Please click here to view a larger version of this figure.
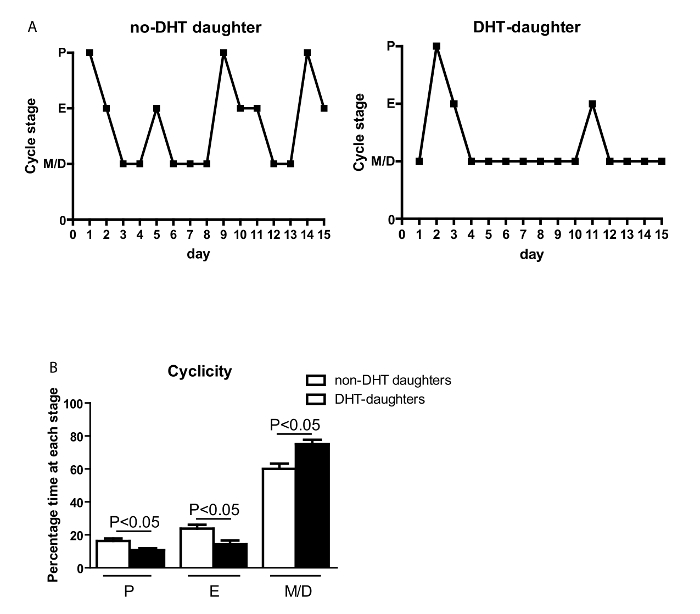
Figure 6: Chronic maternal androgen excess leads to disturbed cyclicity in adult daughters. (A) Representation of estrous cyclicity of female offspring. (B) The percentage of time spent at each estrous stage (Y-axis) during 15 days (X-axis) measured by cytological examination of vaginal cells. Values are mean ±S.E.M. N = 5-9 per group. The estrous cycle stage (Y-axis). M/D: met/diestrus; P: proestrus; E: estrus. Please click here to view a larger version of this figure.
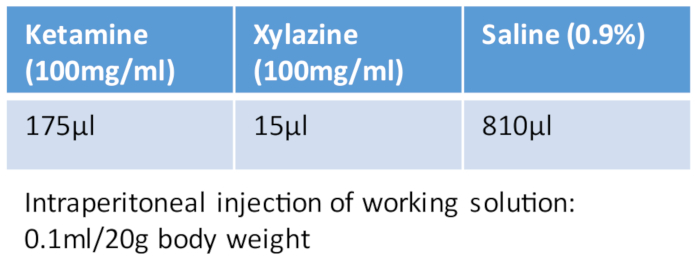
Table 1: Ketamine/xylazine cocktail.
| Crystalline 5α-DHT powder | Sigma-Aldrich | A8380-1G | ||
| Dow Corning Silastic tubing | Fisher Scientific | 11-189-15D | 0.04in/1mm inner diameter x0.085in/2.15mm outer diameter | |
| Medical adhesive silicone | Factor II, InC. | A-100 | ||
| Goggles, lab coats, gloves and masks. | ||||
| 10 µL pipette tips without filter | USA Scientific | 11113700 | ||
| Microscope slide for smear | Fisher Scientific | 12-550-003 | ||
| Diff Quik for staining cells | Fisher Scientific | NC9979740 | ||
| Lancet | Fisher Scientific | NC9416572 | ||
| 3 mL Syring | Becton, Dickinson and Company (BD), | 30985 | ||
| attached needle: 20G | BD | 305176 | ||
| Ruler: any length than 10cm with milimeter scale. | ||||
| Xylazine | Vet one AnnSeA LA, MWI, Boise | NDC13985-704-10 | 100mg/ml | |
| Ketamine Hydrochloride | Hospira, Inc | NDC 0409-2051-05 | 100mg/ml | |
| Surgical staple | AutoClip® System, Fine Science Tool | 12020-00 | ||
| Insulin syringe | BD | 329461 | 1/2 CC, low dose U-100 insulin syringe | |
| Trochar | Innovative Research of America | MP-182 | ||
| Microscope | Carl Zeiss Primo Star | 415500-0010-001 | Germany | |
| Ear punch | Fisher Scientific | 13-812-201 | ||
| Testosterone rat/mouse ELISA kit | IBL | B79174 | ||
| DHT ELISA kit | Alpha Diagnostic International | 1940 | ||
| One touch ultra glucometer | Life Scan, Inc. | |||
| One touch ultra test stripes | Life Scan, Inc. | |||
| Eppendorf tube | Fisher Scientific | 05-402-18 | ||
| Razor blade | Fisher Scientific | 12-640 | ||
| Clidox | Fisher Scientific | NC0089321 | ||
| surgical underpad | Fisher Scientific | 50587953 |
|
|
| Betadine Antiseptic Solution | Walgreens | |||
| 3M Vetbond (n-butyl cyanoacrylate) | 3M Science. Applied to Life | |||
| Animal tattoo ink paste | Ketchum manufacturing Inc. | Brockville, Ontario, Canada | ||
| Scale | Ohaus Corporation | HH120D | Pine Brook, NJ | |
| Electronic digital caliper | NEIKO Tools USA | 01407A | available from Amazon |
