Las mitocondrias cumplen con la provisión clave de energía y son un orgánulo compartimentado que contribuye a procesos bioenergéticos y metabólicos celulares esenciales como el anabolismo de nucleótidos, lípidos y aminoácidos, la biogénesis del grupo hierro-azufre y están implicadas en la señalización como la muerte celular controlada 1,2,3 . La bioenergética mitocondrial a través de la fosforilación oxidativa contribuye a casi todos los procesos celulares dentro de la célula, y en consecuencia, las disfunciones mitocondriales de origen primario o secundario se asocian con un amplio espectro de condiciones de enfermedad 4,5. La disfunción mitocondrial no sólo implica alteraciones en la estructura o densidad mitocondrial sino también en la calidad y regulación del sistema respiratorio6. Este elemento cualitativo abarca el control del sustrato, las características de acoplamiento, las modificaciones post-traduccionales, la dinámica cristae y los supercomplejos respiratorios 7,8. Por lo tanto, el análisis preciso de la bioenergética mitocondrial para enfoques experimentales y diagnósticos para evaluar el metabolismo energético de la célula es importante en la salud y la enfermedad.
La fosforilación oxidativa mitocondrial (OXPHOS) es una secuencia de reacciones dentro del sistema respiratorio o sistema de transferencia de electrones (ETS) para la generación de energía celular a través del trifosfato de adenosina (ATP)9. El paso multienzimático para aprovechar la energía del flujo de electrones a través de los complejos I y II al complejo IV genera un gradiente electroquímico de protones a través de la membrana mitocondrial interna, posteriormente utilizado para la fosforilación de adenosina difosfato (ADP) a ATP a través del complejo V (F1FO ATP sintasa) (Figura 1A).
En primer lugar, se generan portadores de dos electrones durante el ciclo tricarboxílico (TCA), la glucólisis y la oxidación de piruvatos: nicotinamida adenina dinucleótido (NADH) y dihidroflavina adenina dinucleótido (FADH2). El NADH se oxida en el complejo I (NADH deshidrogenasa), durante el cual dos electrones se transfieren a la coenzima Q (la quinona se reduce a quinol), mientras que los protones se bombean al espacio intermembrana (IMS). En segundo lugar, el complejo II (succinato deshidrogenasa) oxida FADH2 y alimenta los electrones a la coenzima Q sin bombear protones. En tercer lugar, en el complejo III (citocromo c oxidorreductasa), los electrones de la coenzima Q se transfieren al citocromo c, mientras que los protones se bombean al IMS. Cuarto, el citocromo c transfiere los electrones al complejo IV (citocromo c oxidasa), el complejo final para bombear protones, y donde el oxígeno funciona como un aceptor de electrones para asimilar protones, formando finalmente agua. Es este oxígeno el que consumen las mitocondrias el que se puede medir mediante un oxígrafo. Finalmente, los protones generados a partir de los complejos I, III y IV se utilizan para rotar el complejo V, generando así ATP9.
Es importante destacar que la transferencia de electrones ocurre no solo de manera lineal, también denotada como la cadena de transporte de electrones. En cambio, los electrones se pueden transferir al grupo de coenzima Q a través de múltiples vías respiratorias y facilitar el flujo convergente de electrones. Los sustratos de NADH y succinato, por ejemplo, pueden entrar a través del complejo I y el complejo II, respectivamente. Los electrones de la oxidación de ácidos grasos se pueden donar a través del complejo de flavoproteína de transferencia de electrones. De hecho, un análisis exhaustivo de OXPHOS requiere un enfoque holístico con sustratos de combustible apropiados (Figura 1A).
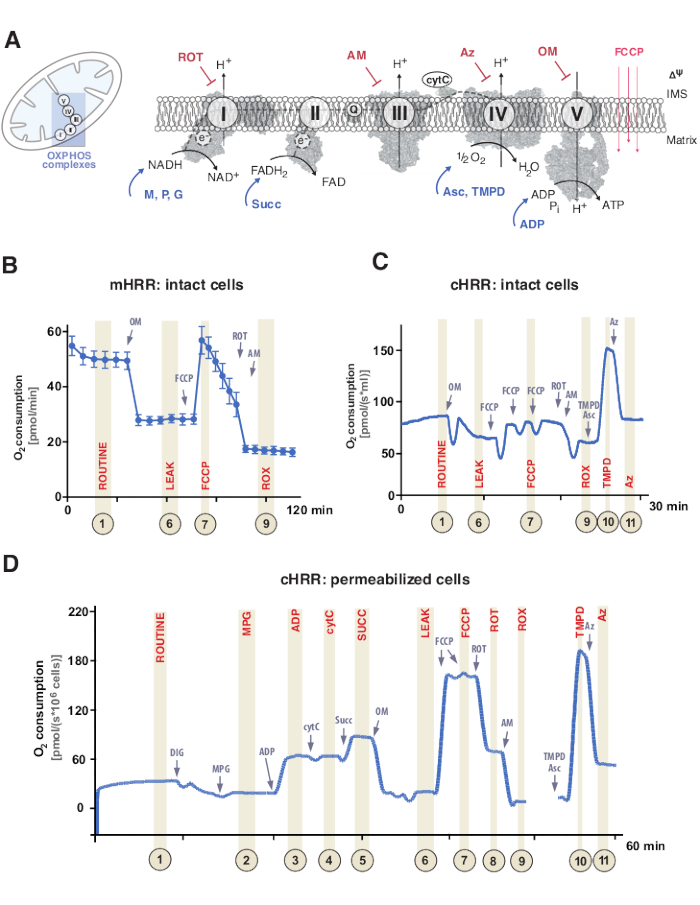
Figura 1: Fosforilación oxidativa mitocondrial y sustrato específico y protocolos inhibidores. (A) Mitocondria y esquema del sistema de transferencia de electrones (CI-CIV) y mitocondrial F1F0 ATP sintasa (CV). Todas las estructuras son de PDB. Las figuras solo representan sustratos e inhibidores descritos en este estudio). (B) Traza de muestra de flujo de oxígeno en células HEK293 intactas utilizando el protocolo estándar en un dispositivo mHRR. (C) Traza de muestra de flujo de oxígeno en células HEK293 intactas utilizando el protocolo estándar en un dispositivo cHRR. (D) Muestra de rastro de flujo de oxígeno en fibroblastos humanos permeabilizados de un donante sano con el protocolo SUIT respectivo. Abreviaturas: 1 = Respiración rutinaria de células intactas; 2 = Estado 2; 3 = Estado 3(I); 4 = Estado 3(I) con cytC; 5 = Estado 3 (I+II); 6 = Fuga (OM); 7 = capacidad del RCDE; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Rotenona, AM = Antimicina, ATP = Trifosfato de adenosina, Az = Azida, OM = Oligomicina, FCCP = Cianuro de carbonilo p-trifluoro-metoxifenil-hidrazona; Asc = Ascorbato, TMPD = N,N,N′,N′-tetrametil-p-fenilendiamina, Succ = Succinato, M = Malato, P = Piruvato, ADP = Adenosina difosfato, NAD = Nicotinamida adenina dinucleótido, IMS = Espacio intermembrana, FAD = Flavina adenina dinucleótido. Haga clic aquí para ver una versión más grande de esta figura.
El análisis de la capacidad mitocondrial de OXPHOS utilizando HRR se ha convertido en un método bioquímico instrumental de valor diagnóstico no solo para defectos mitocondriales primarios10,11, sino que se extiende a todos los demás ámbitos de la biología, como el cáncer y el envejecimiento12. La HRR permite la determinación de la respiración celular mediante el análisis de la capacidad mitocondrial de OXPHOS, que refleja directamente la deficiencia del complejo respiratorio mitocondrial individual o combinada, e indirectamente se asocia con disfunción celular y metabolismo energético alterado9. Metodológicamente, las mediciones de la respiración se realizan utilizando células, tejidos o mitocondrias aisladas 11,13,14, con material congelado solo parcialmente adecuado15,16. Se ha demostrado que el tejido congelado tiene un ETS intacto con una estabilidad supercompleja mantenida15. Por lo tanto, a diferencia de los intermedios tradicionales de TCA, los sustratos respectivos se introducen directamente en el ETS. Sin embargo, el acoplamiento entre el ETS y la síntesis de ATP se pierde a medida que la integridad de la membrana se ve comprometida por el daño por congelación (formación de cristales de hielo).
Los experimentos de respiración normalmente tienen lugar a una temperatura fisiológica de 37 °C para endotermas en células o tejidos no permeabilizados o permeabilizados. Mientras que el primero considera el contexto metabólico citosólico, el segundo proporciona la contribución energética de los complejos OXPHOS individuales y la ATPasa a través de la adición de sustratos específicos (e inhibidores). La secuencia y la variación de sustratos e inhibidores han llevado al desarrollo de una amplia gama de protocolos SUIT17 y ensayos18 para abordar diversas cuestiones científicas de la función oxphos (revisados bajo12). El protocolo básico de la respiración celular evalúa cuatro estados diferentes: i) respiración de rutina: la respiración en un medio respiratorio respectivo sin ninguna adición de sustratos o inhibidores que consuman sino sustratos endógenos. Este estado puede revelar OXPHOS general o defectos respiratorios inducidos secundariamente causados, por ejemplo, por perfiles de metabolitos alterados. A continuación, la adición del inhibidor de la ATPasa oligomicina revela la permeabilidad de la membrana mitocondrial interna a los protones, definida como ii) respiración por fuga. La titulación posterior de un protonóforo como el cianuro de carbonilo no acoplado p-trifluoro-metoxifenil-hidrazona (FCCP) permite determinar el estado en el que la capacidad de ETS es máxima en un modo de circuito de protones transmembrana abierta, definido como iii) respiración desacoplada. Es importante destacar que un estado desacoplado también puede ocurrir por intervenciones experimentales a través de un daño mecánico excesivo a las membranas mitocondriales. Por el contrario, el estado no acoplado se refiere al desacoplamiento respiratorio por un mecanismo intrínseco que está fisiológicamente controlado. Finalmente, la inhibición completa del ETS mediante la adición del inhibidor del complejo III antimicina y el inhibidor del complejo I rotenona determina el consumo residual de oxígeno (ROX) de procesos no mitocondriales que consumen oxígeno (Figura 1A-C).
La bioenergética mitocondrial consta de cinco estados respiratorios distintos19,20. La respiración de estado 1 es sin sustratos adicionales o ADP, excepto por lo que está disponible endógenamente. Después de la adición de ADP, pero aún así, sin sustratos, se logra la respiración del estado 2. Cuando se agregan sustratos, lo que permite la transferencia de electrones y la síntesis de ATP, se alcanza la respiración del estado 3. En este estado, la capacidad de OXPHOS se puede definir a concentraciones saturadoras de ADP, fosfato inorgánico, oxígeno, sustratos ligados a NADH y succinato. La respiración de estado 4 o respiración LEAK se puede definir como un estado sin ADP o ATP sintasas inhibidas químicamente mientras que tiene sustratos suficientes. Por último, cuando todo el oxígeno se agota (anóxico) en un entorno de cámara cerrada, se observa la respiración de estado 5.
Existen varios métodos para evaluar los estados de energía celular14 con dos dispositivos que dominan la evaluación actual en tiempo real de OXPHOS a través del análisis del consumo de oxígeno, medido como la función de la disminución de oxígeno a lo largo del tiempo en un sistema de cámara cerrada con diferente aplicabilidad dependiendo del modelo experimental y la pregunta de investigación: el respirómetro de alta resolución Oroboros 2k y el analizador de flujo extracelular Seahorse XF. Ambos dispositivos registran las tasas de consumo de oxígeno como una disminución en picomoles (pmol) de oxígeno (O2) por segundo como un valor absoluto dentro de la cámara o microplaca del pozo. El consumo específico de oxígeno por masa se obtiene normalizando el consumo de oxígeno respectivo en una receta tampón específica por número de células (millones), peso tisular (mg) o cantidad de proteína.
El O2k (Oroboros Instruments) es un sistema cerrado de dos cámaras equipado con un sensor polarográfico de oxígeno (abreviado como respirómetro de alta resolución basado en cámara: cHRR). Cada cámara experimental contiene 2 ml de líquido que se mantiene homogéneo por agitadores magnéticos. El sensor polarográfico de oxígeno utiliza un enfoque amperométrico para medir el oxígeno: contiene un cátodo de oro, un ánodo de cloruro de plata / plata y, entre una solución KCI, crea una celda electroquímica sobre la cual se aplica un voltaje (0.8 V). El oxígeno del medio de ensayo se difunde a través de una membrana de etileno propileno fluorado de 25 μm (O 2-permeable) y sufre una reducción en el cátodo, produciendo peróxido de hidrógeno. En el ánodo, la plata es oxidada por peróxido de hidrógeno, generando una corriente eléctrica. Esta corriente eléctrica (amperio) está linealmente relacionada con la presión parcial de oxígeno. La presión parcial de oxígeno y el factor de solubilidad de oxígeno del medio de ensayo se utilizan para calcular la concentración de oxígeno. Dado que la presión parcial de oxígeno depende de la temperatura experimental y las mediciones polarográficas son sensibles a la temperatura, las fluctuaciones de temperatura necesitan una regulación precisa (±0,002 °C) mediante un bloque de calentamiento Peltier. La temperatura se puede controlar dentro de un rango de 4 ° C y 47 ° C.
El analizador de flujo extracelular Seahorse XF (Agilent) es un sistema basado en placas con formato de microplaca de 24 o 96 pocillos en el que tres electrodos de fluorescencia miden el consumo de oxígeno a lo largo del tiempo en cada pozo (abreviado como respirómetro de alta resolución basado en microplacas: mHRR). Un máximo de cuatro puertos en el cartucho de ensayo están disponibles para la inyección automatizada durante el ensayo. Un ensayo contiene múltiples ciclos, cada uno con tres fases: 1) mezcla, 2) espera y 3) medición. Durante la fase de medición, las sondas del sensor se bajan a la microplaca creando una cámara temporalmente cerrada que contiene un volumen de 7-10 μL para medir la luz emitida. Esta luz es emitida por fluoróforos incrustados en polímeros en la punta de las sondas del sensor, que detectan O2 en función del enfriamiento de fosforescencia. La intensidad de la señal de fluorescencia es proporcional a O2 e influenciada por la temperatura del sensor y el medio de ensayo. Por lo tanto, la estimación precisa del oxígeno requiere un enfoque relativo con un pozo de fondo sin ninguna muestra. La restauración de la concentración de oxígeno ocurre durante la fase de mezcla cuando el sensor se mueve hacia arriba y hacia abajo para mezclar el volumen por encima de la cámara temporal. Cada ciclo calcula una tasa de consumo de oxígeno. La temperatura se puede controlar dentro de un rango de 16 ° C y 42 ° C.
HrR es el estándar de oro para evaluar la bioenergética celular en enfermedades primarias y asociadas a las mitocondrias y el metabolismo celular general. En este estudio, se proporcionan protocolos básicos para la HRR para evaluar la función de OXPHOS en células y tejidos.
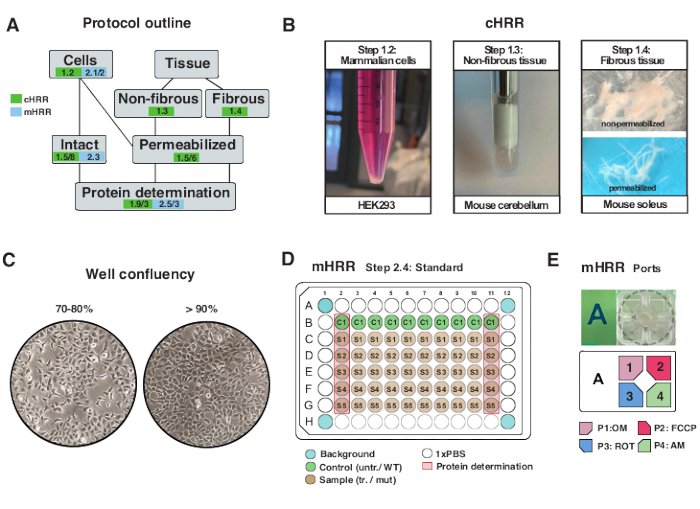
Figura 2: Flujo de trabajo para preparaciones celulares y tisulares para la RCH, y preparación celular para la respirometría de RMH. (A) Esquema de los protocolos proporcionados. (B) Células de mamíferos (paso 1.2): pellet HEK293 igual a 3 x 106 celdas (panel izquierdo). Tejido no fibroso (paso 1.3): Preparación de lisado de cerebelo murino en alfarero de teflón de 2 ml (panel medio). Permeabilización del músculo esquelético inducida por saponina (paso 1.4) panel derecho) para la respirometría de RCH. (C) Diseño estándar de siembra de microplacas (paso 2.4) y verificación de confluencia para el análisis de células eucariotas (HEK293) para la respirometría mHRR. (D, E) Esquema de carga del puerto de inyección para respirometría mHRR (paso 2.4). Haga clic aquí para ver una versión más grande de esta figura.
