Большое количество событий подвижности в эукариотических клетках опосредовано функцией молекулярных моторных белков. Эти двигатели движутся вдоль цитоскелетных нитей, актиновых нитей и микротрубочек (МТ) и преобразуют химическую энергию гидролиза АТФ в кинетические и механические силы, необходимые для управления биологической подвижностью внутри клеток. S. cerevisiae Cin8 на основе MT представляет собой биполярный гомотетрамерный моторный белок кинезин-5, который сшивает и скользит веретено MTs друг отдруга 1. Cin8 выполняет важнейшие функции при митозе, в веретенообразном сборе 2,3,4 и удлинении шпинделя при анафазе 5,6,7. Ранее было продемонстрировано, что Cin8 является двунаправленным двигателем, который переключает направленность в разных экспериментальных условиях. Например, в условиях высокой ионной прочности одиночные двигатели Cin8 движутся к минусовому концу MT, в то время как в кластерах, в многомоторных скользящих анализах MT и между противопараллельными MT двигатели Cin8 движутся в основном к плюсовым концам MTs 8,9,10,11,12 . Эти результаты были весьма неожиданными по нескольким причинам. Во-первых, Cin8 несет свой каталитический двигательный домен на амино-конце, и ранее считалось, что такие двигатели направлены исключительно плюс-конец, тогда как Cin8 был показан как минус-конец, направленный на уровень одной молекулы. Во-вторых, кинезиновые двигатели считались однонаправленными, либо минусовыми, либо плюс-концевыми, тогда как Cin8 был показан двунаправленным, в зависимости от условий эксперимента. Наконец, из-за ориентации МТ на митотическом шпинделе классическая роль двигателей кинезин-5 в разделении полюсов шпинделя при сборке шпинделя и анафазы B может быть объяснена только их направленной подвижностью на МТ, которые они сшивают 1,13. После первых сообщений о двунаправленности Cin8 было продемонстрировано, что несколько других кинезиновых двигателей являются двунаправленными 14,15,16, что указывает на то, что двунаправленная подвижность кинезиновых двигателей может быть более распространенной, чем считалось ранее.
Ранее сообщалось, что в клетках Cin8 также движется двунаправленным образом8, поддерживая представление о том, что двунаправленная подвижность некоторых двигателей кинезина-5 важна для их внутриклеточных функций. Кроме того, поскольку три двигателя кинезин-5, которые, как сообщалось, являются двунаправленными, получены из грибковых клеток, в таких клетках10 недавно была предложена возможная роль двунаправленности двигателей кинезин-5. Согласно этой модели, при закрытом митозе грибковых клеток, где ядерная оболочка не разрушается во время митоза, двигатели кинезина-5 обеспечивают начальную силу, которая разделяет полюса шпинделя друг от друга до сборки шпинделя. Для выполнения этой задачи до разделения полюсов шпинделя двигатели кинезин-5 локализуются вблизи полюсов шпинделя своей направленной подвижностью на одиночных ядерных МТ. Оказавшись в этом положении, двигатели кинезин-5 группируются, переключают направленность, захватывают и сшивают МТ с соседних полюсов шпинделя. Впоследствии двигатели кинезин-5 обеспечивают начальное разделение полюсов за счет плюсовой направленной подвижности на МТ, которые они сшивают. В соответствии с этой моделью как направленная подвижность на одиночных МТ, так и направленная подвижность на сшитых МТ во время антипараллельного скольжения необходимы для выполнения грибковых двигателей кинезин-5 для выполнения своих ролей в сборке шпинделя 1,13.
Общей целью описанного способа является получение высокочистого грибка GFP-меченого кинезина-5 Cin8 и выполнение анализа подвижности одной молекулы (рисунок 1) при отдельном анализе подвижности отдельных молекул и кластеров Cin8. Разделение между отдельными молекулами и кластерами важно, поскольку одним из факторов, которые, как было продемонстрировано, влияют на направленность Cin8, является его накопление в кластерах на MTs10,12. Альтернативные анализы подвижности, такие как поверхностное скольжение MT и скользящие анализы MT, не дают информации об активности одиночных моторных белков17,18. Надежные одномолекулярные методы анализа подвижности и анализа, описанные здесь, были успешно применены для характеристики различных аспектов двигателей кинезин-5, Cin8 и Kip1 10,11,12,14,19,20.
Здесь представлен подробный протокол для сверхэкспрессии и очистки Cin8, полимеризации МТ и анализа подвижности одной молекулы. Кроме того, описаны анализы для дифференциации отдельных молекул и кластеров Cin8, а также для определения одиночных моторных и кластерных скоростей с помощью анализа среднего смещения (MD) и среднего квадратного смещения (MSD). Этот протокол призван помочь исследователям визуализировать все этапы процедур и помочь в устранении неполадок этого типа анализов.
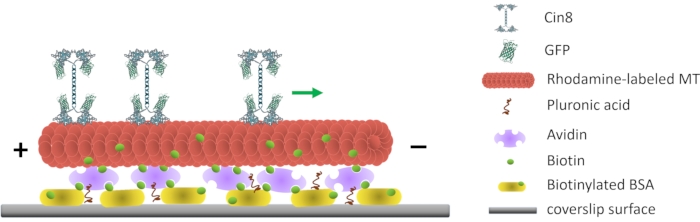
Рисунок 1: Схематическое представление анализа подвижности одной молекулы. Биотинилированные флуоресцентные МТ прикреплены к поверхности стекла, покрытой авидином, который взаимодействует с поверхностно присоединенным биотинилированным BSA. Зеленая стрелка представляет направление движения отдельных молекул Cin8 в условиях высокой ионной прочности. +/- представляют полярность MT. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
