


Summary
Dit artikel beschrijft een open-source digitaal beeldcorrelatiealgoritme voor het meten van lokale 2D-weefselspanningen in peesexplantaten. De nauwkeurigheid van de techniek is gevalideerd met behulp van meerdere technieken en is beschikbaar voor openbaar gebruik.
Abstract
Er is veel wetenschappelijke interesse in het begrijpen van de stammen die peescellen in situ ervaren en hoe deze stammen weefselremodellering beïnvloeden. Op basis van deze interesse zijn verschillende analytische technieken ontwikkeld om lokale weefselspanningen in peesexplantaten te meten tijdens het laden. In verschillende gevallen zijn de nauwkeurigheid en gevoeligheid van deze technieken echter niet gerapporteerd en geen van de algoritmen is openbaar beschikbaar. Dit heeft het moeilijk gemaakt voor de meer wijdverspreide meting van lokale weefselspanningen in peesexplantaten. Daarom was het doel van dit artikel om een gevalideerde analysetool te creëren voor het meten van lokale weefselspanningen in peesexplantaten die direct beschikbaar en gemakkelijk te gebruiken is. In het bijzonder werd een openbaar beschikbaar augmented-Lagrangian digital image correlation (ALDIC) -algoritme aangepast voor het meten van 2D-stammen door de verplaatsingen van celkernen in achillespezen van muizen onder uniaxiale spanning te volgen. Bovendien werd de nauwkeurigheid van de berekende stammen gevalideerd door digitaal getransformeerde beelden te analyseren en door de stammen te vergelijken met waarden die zijn bepaald met behulp van een onafhankelijke techniek (d.w.z. gefotobleekte lijnen). Ten slotte werd een techniek in het algoritme opgenomen om het referentiebeeld te reconstrueren met behulp van het berekende verplaatsingsveld, dat kan worden gebruikt om de nauwkeurigheid van het algoritme te beoordelen bij afwezigheid van bekende rekwaarden of een secundaire meettechniek. Het algoritme is in staat om stammen tot 0,1 te meten met een nauwkeurigheid van 0,00015. De techniek voor het vergelijken van een gereconstrueerd referentiebeeld met het werkelijke referentiebeeld identificeerde met succes monsters met onjuiste gegevens en gaf aan dat, in monsters met goede gegevens, ongeveer 85% van het verplaatsingsveld nauwkeurig was. Ten slotte waren de stammen gemeten in achillespezen van muizen consistent met de eerdere literatuur. Daarom is dit algoritme een zeer nuttig en aanpasbaar hulpmiddel voor het nauwkeurig meten van lokale weefselspanningen in pezen.
Introduction
Pezen zijn mechanosensitieve weefsels die zich aanpassen en degenereren als reactie op mechanische belasting 1,2,3,4. Vanwege de rol die mechanische stimuli spelen in de biologie van peescellen, is er een grote interesse in het begrijpen van de spanningen die peescellen ervaren in de inheemse weefselomgeving tijdens het laden. Er zijn verschillende experimentele en analytische technieken ontwikkeld om lokale weefselspanningen in pezen te meten. Deze omvatten 2D / 3D digitale beeldcorrelatie (DIC) analyses van oppervlaktespanningen met behulp van spikkelpatronen of gebleekte lijnen (PBL's) 5,6,7,8, meting van de veranderingen in de centroïde-tot-centroïde afstand van individuele kernen in het weefsel 9,10, en een recente full-field 3D DIC-methode die rekening houdt met out-of-plane beweging en 3D-vervormingen 11 . De nauwkeurigheid en gevoeligheid van deze technieken zijn echter slechts in enkele gevallen gemeld en geen van deze technieken is openbaar gemaakt, wat de wijdverbreide toepassing en het gebruik van deze technieken moeilijk maakt.
Het doel van dit werk was om een gevalideerd analyse-instrument te creëren voor het meten van lokale weefselspanningen in peesexplantaten dat direct beschikbaar en gemakkelijk te gebruiken is. De gekozen methode is gebaseerd op een openbaar beschikbaar augmented-Lagrangian digital image correlation (ALDIC) algoritme geschreven in MATLAB dat is ontwikkeld door Yang en Bhattacharya12. Dit algoritme werd aangepast voor het analyseren van peesmonsters en gevalideerd door het toe te passen op digitaal getransformeerde beelden en door de stammen gemeten in werkelijke peesmonsters te vergelijken met de resultaten verkregen uit gefotobleekte lijnen. Bovendien werd extra functionaliteit in het algoritme geïmplementeerd om de nauwkeurigheid van het berekende verplaatsingsveld te bevestigen, zelfs als er geen bekende rekwaarden of een secundaire meettechniek waren. Daarom is dit algoritme een zeer nuttig en aanpasbaar hulpmiddel voor het nauwkeurig meten van lokale 2D-weefselspanningen in pezen.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
Deze studie werd goedgekeurd door de Pennsylvania State University Institutional Animal Care and Use Committee.
1. Weefselvoorbereiding
- Oogst voor dit protocol de achillespezen van 2-4 maanden oude mannelijke C57BL / 6-muizen.
OPMERKING: Verschillende pezen of ligamenten van muizen of andere kleine dieren kunnen ook worden gebruikt.- Maak een incisie in de huid oppervlakkig naar de achillespees om de plantarispees en het omliggende bindweefsel bloot te leggen. Verwijder ze vervolgens met een chirurgisch mes.
- Scheid de blootgestelde soleus- en gastrocnemiusspieren van de achterpoot en schraap ze voorzichtig van de achillespees met het chirurgische mes
- Scheid de calcaneus van de rest van de voet met een snijwielbevestiging op een roterend gereedschap.
- Kleur het weefsel in 1,5 ml van een 5 μg/ml oplossing van 5-(4,6-dichloortriazinyl) aminofluoresceïne (DTAF) en 0,1 M natriumbicarbonaatbuffer gedurende 20 minuten op een roterende mixer bij kamertemperatuur. Deze oplossing kleurt eiwitten (bijv. Extracellulaire matrix) in het weefsel.
OPMERKING: Tijdens deze periode van 20 minuten moet stap 1.3 worden voltooid. - Bereid een 1:1.000 oplossing van DRAQ5 in fosfaat-gebufferde zoutoplossing (PBS) om de kernen te kleuren. Gebruik een vortexmixer om de oplossing te homogeniseren.
- Breng na de incubatietijd van 20 minuten in stap 1.2 het weefsel over van de DTAF-oplossing naar de DRAQ5-oplossing en incubeer gedurende 10 minuten in een donkere ruimte bij kamertemperatuur.
2. Peesbelasting en beeldacquisitie
OPMERKING: Dit protocol vereist een trekapparaat dat bovenop een confocale microscoop kan worden gemonteerd. Voor deze studie werd het microtensile apparaat beschreven door Peterson en Szczesny13 gebruikt.
- Plaats de pees in de grepen van de trekbelasting. Voordat u de handgrepen in de laadinrichting monteert, gebruikt u digitale remklauwen om de afstand tussen het calcaneus-hulpstuk en de tegenovergestelde greep te meten. Deze afstand is de lengte van de peesmeter.
- U kunt ook de grepen in de laadinrichting monteren voordat u de pees inbrengt en in contact duwen om de positie van de motor zonder verplaatsing te bepalen. De verplaatsing van de motoren na het inbrengen van de pees zou een potentieel nauwkeurigere grip-to-grip meterlengte kunnen bieden.
- Monteer de grepen in het laadapparaat, dat PBS bevat om de weefselhydratatie te behouden. Lijn de pees zo goed mogelijk uit met de x-as of y-as van de microscoopbeelden, zodat de x-rek- en y-rekuitgangen van het algoritme overeenkomen met de peesassen.
OPMERKING: In deze studie werden de pezen uitgelijnd met de x-as. Als het niet mogelijk is om de pees perfect uit te lijnen met de beeldassen, dan kunnen de x-strain en y-strain outputs van het algoritme worden getransformeerd om uit te lijnen met de longitudinale / loodrechte assen van de pees met behulp van standaard spanningstransformatievergelijkingen14. - Laad de pees voor met 1 g spanning en pas, indien gewenst, cyclische belasting toe om het monster voor te bereiden. In dit protocol werd geen voorconditionering gebruikt, omdat het doel van de studie was om de gemeten lokale weefselstammen te valideren in plaats van de eigenschappen van het weefselmateriaal te meten. Als er interesse is in het meten van de materiaaleigenschappen op macroschaal, die afhankelijk zijn van de laadgeschiedenis, dan is voorconditionering aan te bevelen. Na het conditioneren en herstellen, opnieuw aanbrengen van een voorspanning van 1 g.
- Indien gewenst, fotobleekt een set van vier lijnen met een onderlinge afstand van 80 μm in het middengebied van het weefsel (zie Peterson en Szczesny13 voor meer details).
OPMERKING: De gefotobleekte lijnen werden gebruikt om de metingen van het ALDIC-algoritme te valideren en zijn niet nodig voor het uitvoeren van de ALDIC zelf. Het aantal en de afstand van de lijnen kunnen worden aangepast en de locatie van de lijnen moet worden gekozen om artefacten in de steekproef te voorkomen die de helderheid van de lijn zouden verminderen. - Herhaal de fotobleekprocedure aan de linker- en rechteruiteinden van het weefsel in de buurt van de grepen.
- Verkrijg met behulp van de confocale microscoop volumetrische beelden (x,y: 1,25 μm/pixel, z: 2,5 μm/pixel) van de DTAF- en DRAQ5-fluorescentie bij 1 g voorspanning.
- Voer een rekhelling uit met een spanning van 0,5% /s tot 2%. Merk op dat de reksnelheid en de incrementele spanningsgrootte kunnen worden aangepast.
- Laat het weefsel 10 minuten stressen.
OPMERKING: De duur van de ontspanning van de spanning moet zodanig worden gekozen dat het monster tijdens de beeldacquisitie onder een ongeveer quasistatische belasting staat. Om te bepalen of de duur van de stressontspanning acceptabel is, bepaalt u de helling van de kracht-tijdcurve tijdens de laatste minuut van de stressontspanning (aanvullende figuur 1) en vermenigvuldigt u deze helling met de totale beeldduur. In deze studie veranderde de kracht die werd uitgeoefend bij de grootste rektoename nooit met meer dan 5%. - Neem nog een volumetrisch beeld van het weefsel na vervorming.
- Herhaal stap 2.7-2.9 totdat de gewenste laatste stam is bereikt. In dit artikel is gekozen voor een uiteindelijke rekwaarde van 12%.
3. Beeldverwerking
- Gebruik ImageJ of Fiji om maximale z-projecties te maken van elk volumetrische beeld van het DRAQ5 (nucleaire) kanaal. Dit zal dienen als de 2D-gespikkelde beelden voor de ALDIC.
- Sla de z-projecties met maximale intensiteit op als .tiff bestanden en geef ze een naam volgens de volgende naamgevingsconventie.
- Gebruik een getal als eerste teken van de afbeeldingsnaam.
- Laat het nummer overeenkomen met de volgorde waarin de beelden tijdens de stamanalyse worden bekeken. De eerste afbeelding moet bijvoorbeeld met één beginnen en de tweede afbeelding moet met twee beginnen. Er kunnen verschillende getallen worden gekozen, maar ze moeten sequentieel toenemen. Een voorbeeld van een naamgevingsconventie is als volgt: "0_Experiment1_MaxZProjection".
- Sla alle hernoemde max-intensity z-projecties op in een map.
4. Fotogebleekte lijnanalysecode installatie en toepassing
OPMERKING: Deze stappen zijn alleen nodig als het gewenst is om de nauwkeurigheid van het ALDIC-algoritme te bevestigen met behulp van gebleekte lijnen. De code berekent de lokale weefselstam als de gemiddelde genormaliseerde verandering in afstand tussen elke gefotobleekte lijn binnen de gefotobleekte lijnset. In deze studie werden de gemiddelde lokale waarden vervolgens gemiddeld over alle gefotobleekte lijnsets (d.w.z. in het midden en de linker / rechteruiteinden) om een enkele gemiddelde lokale weefselstamwaarde voor elk monster te bepalen. Deze waarde werd vervolgens gebruikt om de nauwkeurigheid van het ALDIC-algoritme te schatten.
- Download de map "PGO-code" van GitHub (https://github.com/Szczesnytendon/TendonStrainCalc) en verplaats alle inhoud naar de werkmap in MATLAB.
- Open het MATLAB-script "Micro_Mech_Template.m".
- Druk op Uitvoeren en selecteer een van de afbeeldingsbestanden met de volumetrische afbeeldingen. De volumetrische afbeeldingen kunnen een van de volgende bestandstypen zijn: .lsm, .tiff, .nd2.
- De software laadt automatisch alle afbeeldingen in de map en geeft een geprojecteerd beeld van de referentievolumetrische afbeelding weer. Wanneer u hierom wordt gevraagd, klikt u met de linkermuisknop om meerpuntslijnen te maken die de linker- en rechterkant van het voorbeeld volgen. Klik met de rechtermuisknop om een regel te beëindigen. Zodra de invoer is verwerkt en de randen correct zijn, drukt u op OK om het resultaat te accepteren.
- Teken een willekeurige diagonale lijn over het voorbeeld als referentielijn wanneer daarom wordt gevraagd.
- Voer het aantal gemaakte gefotobleekte lijnen in en volg de gebleekte lijnen met meerpuntslijnen.
- Als het resultaat acceptabel is, accepteer het dan. Als het resultaat onjuist is, pas het dan aan en verwerk het opnieuw.
- Herhaal stap 4.2 voor alle afbeeldingen en verplaats alle afbeeldingen van getraceerde lijnen naar één map.
- Open het script "Micro_Mech_Strain.m".
- Druk op Uitvoeren om de code uit te voeren en selecteer een van de opgeslagen afbeeldingen waar de gefotobleekte lijnen worden overgetrokken.
- Controleer of de geselecteerde begeleidende afbeeldingen correct zijn zodra de afbeelding is geselecteerd door op OK te drukken.
5. Digitaal getransformeerde afbeeldingen maken
OPMERKING: Deze stappen zijn alleen nodig als het gewenst is om de nauwkeurigheid van het ALDIC-algoritme te bevestigen met behulp van digitaal getransformeerde afbeeldingen. Deze beelden simuleren homogene 2D-spanningsvelden van een bekende omvang door het referentiebeeld kunstmatig te transformeren.
- Download de code "Digital_strain.m" van GitHub (https://github.com/Szczesnytendon/TendonStrainCalc).
- Open de code en voer deze uit.
- Wanneer u hierom wordt gevraagd, voegt u de gewenste waarden in voor de maximaal toegepaste stam, de toegepaste stamtoename en de verhouding van Poisson. Druk op OK.
OPMERKING: Voor dit experiment was de maximaal toegepaste stam 0,1 (10%), de toegepaste stamtoename was 0,02 (2%) en een Poisson-verhouding van 1 werd gebruikt, wat consistent is met experimentele gegevens van peestrektesten15,16. De code maakt gebruik van de ingesloten MATLAB-functie imwarp en de invoerwaarden (bijv. rekstappen, Poisson's ratio) om de digitaal getransformeerde afbeeldingen te maken. - Selecteer desgevraagd de niet-gevormde referentieafbeelding.
- Voor elke stamtoename wordt een overlay van de referentieafbeelding en de getransformeerde afbeelding weergegeven. De getransformeerde afbeelding wordt opgeslagen in de map onder de titel "DigitallyTransformedX%Strain", waarbij X de stamtoename is.
6. Rekberekening en validatiecode installatie en toepassing
- Download de map "Strain Calculation and Validation Code" van GitHub (https://github.com/Szczesnytendon/TendonStrainCalc) en verplaats alle inhoud naar de MATLAB-werkmap
- Installeer een mex C/C++ compiler volgens Yang en Bhattacharya12. De stappen worden hieronder samengevat.
- Controleer MATLAB om te zien of er een mex C/C++ compiler is geïnstalleerd door "mex -setup" te typen in het MATLAB Command Window en op Enter te drukken.
- Als er een fout verschijnt die aangeeft dat een compiler niet wordt ondersteund of aanwezig is, gaat u verder met stap 6.3 en stap 6.4.
- Als er geen fout aanwezig is, gaat u verder met stap 6.5
- Om een mex C/C++ compiler te downloaden, ga naar "https:/tdm-gcc.tdragon.net/", en kies de TDM-gcc compiler.
- Installeer de gedownloade compiler op een bekende locatie.
- Ga terug naar het MATLAB-opdrachtvenster en typ: "setenv("MW_MINGW64_LOC","[Typ hier uw installatiepad]")". Dit kan bijvoorbeeld "setenv("MW_MINGW64_LOC","C:\TDM-GCC-64")" zijn. Als deze opdracht met succes wordt uitgevoerd, is de mex-compiler correct geïnstalleerd.
- Voer het functiescript "main_aldic.m" in en wijzig regel 22 zodat deze overeenkomt met de opdracht die in stap 6.5 is uitgevoerd.
- Open het script "Strain_calc_and_validate.m".
- Druk op Uitvoeren om de afbeeldingsanalyse te starten.
- Wijzig desgevraagd de waarden voor de ALDIC-parameters naar wens.
OPMERKING: De venstergrootte moet 0,25 tot 1 keer de grootte van de subset zijn. Voor meer informatie over de parameterkeuzes raadpleegt u de online gebruikershandleiding: (https://www.researchgate.net/publication/344796296_Augmented_Lagrangian_Digital
_Image_Correlation_AL-DIC_Code_Manual).- In dit onderzoek zijn de volgende waarden gebruikt:
Subset Grootte (pixels): 20
Venstergrootte (pixels): 10
Methode om ALDIC op te lossen: eindig verschil (1)
Parallel computing werd niet gebruikt (1)
Methode om de eerste schatting te berekenen: Multigrid-zoekopdracht op basis van afbeeldingspiramide (0)
- In dit onderzoek zijn de volgende waarden gebruikt:
- Schakel desgevraagd het selectievakje "Ja" in om het algoritme automatisch de gemiddelde waarde, standaarddeviatie en 2D-toewijzingen te laten opslaan voor de gewenste verzameling variabelen (bijvoorbeeld x-stam, y-stam, afschuifspanning, slechte regio's, enz.). Selecteer welke variabelen moeten worden opgeslagen en druk op OK.
- Wanneer u hierom wordt gevraagd, wijzigt u de parameters naar wens.
- In dit experiment werden de volgende waarden gebruikt:
Omringende punten om spanning te berekenen (numP): 12
Correlatiecoëfficiënt voor identificatie van slechte regio's (corr_threshold): 0,5
Subregiogrootte (pixels) voor slechte regioanalyse (subgrootte): 32
- In dit experiment werden de volgende waarden gebruikt:
- Wanneer u hierom wordt gevraagd, selecteert u de map met de hernoemde max-intensity z-projecties. Merk op dat de software automatisch incrementele ALDIC uitvoert om de spanningsvelden van de vervormde afbeeldingen te bepalen. Dat wil zeggen, elke vervormde afbeelding dient als de nieuwe "referentie" -afbeelding voor de volgende vervormde afbeelding. Dit verbetert de nauwkeurigheid van de resultaten (aanvullende figuur 2) in vergelijking met het uitvoeren van cumulatieve ALDIC, waarbij elk vervormd beeld wordt vergeleken met het oorspronkelijke (0% stam) referentiebeeld. Als u een cumulatieve analyse wilt uitvoeren, laadt u de afbeeldingen, maar selecteert u alleen de oorspronkelijke referentieafbeelding en de vervormde afbeelding van belang.
OPMERKING: De normale stam wordt berekend als λ - 1, waarbij λ de weefselrek is. De weefselrek wordt berekend volgens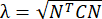 , waarbij N = [1 0]T of [0 1]T voor respectievelijk de x-richting en de y-richting, en C = F T F, waarbij F de vervormingsgradiënt is die wordt berekend met behulp van "numP" -punten rond elke gegevenspuntuitvoer door het ALDIC-algoritme. De schuifspanning wordt berekend als
, waarbij N = [1 0]T of [0 1]T voor respectievelijk de x-richting en de y-richting, en C = F T F, waarbij F de vervormingsgradiënt is die wordt berekend met behulp van "numP" -punten rond elke gegevenspuntuitvoer door het ALDIC-algoritme. De schuifspanning wordt berekend als 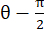 , waarbij
, waarbij 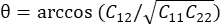 .
. - Wanneer u hierom wordt gevraagd, klikt u met de linkermuisknop om een vierpuntsveelhoek te maken om het gebied te definiëren dat van belang is voor het meten van de stammen. Begin met het punt in de linkerbovenhoek en wijs de volgende punten met de klok mee toe.
OPMERKING: De variabele "Storage" die is opgeslagen in de MATLAB-werkruimte bevat alle waarden voor de gemiddelde x-strain, x-strain standaarddeviatie, gemiddelde y-strain, y-strain standaarddeviatie, gemiddelde shear strain, shear strain standard deviation en percentage bad regions. De slechte regio's worden gedefinieerd volgens de correlatiecoëfficiëntanalyse binnen de regio van belang die is geselecteerd in stap 6.13. De map "NuclearTrackingResults" (die kan worden hernoemd door de regels 555 en 556 aan te passen) slaat alle plots op die zijn opgegeven in stap 6.10. Deze map bevat ook een spreadsheetbestand met de naam "Resultaten", waarin alle gemiddelden en standaardafwijkingen worden opgeslagen die in stap 6.10 zijn gespecificeerd.
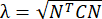 , waarbij N = [1 0]T of [0 1]T voor respectievelijk de x-richting en de y-richting, en C = F T F, waarbij F de vervormingsgradiënt is die wordt berekend met behulp van "numP" -punten rond elke gegevenspuntuitvoer door het ALDIC-algoritme. De schuifspanning wordt berekend als
, waarbij N = [1 0]T of [0 1]T voor respectievelijk de x-richting en de y-richting, en C = F T F, waarbij F de vervormingsgradiënt is die wordt berekend met behulp van "numP" -punten rond elke gegevenspuntuitvoer door het ALDIC-algoritme. De schuifspanning wordt berekend als 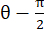 , waarbij
, waarbij 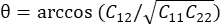 .
.Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Voorafgaand aan het analyseren van de stamvelden in echte weefselmonsters, werd het ALDIC-protocol eerst gevalideerd met behulp van digitaal gespannen / getransformeerde beelden van kernen in achillespezen van muizen. In het bijzonder werden de beelden getransformeerd om digitaal uniforme stammen te produceren in de x-richting van 2%, 4%, 6%, 8% en 10% spanning met een gesimuleerde Poisson-verhouding van 115,16. De nauwkeurigheid van het ALDIC-algoritme werd vervolgens beoordeeld door de gemiddelde berekende stamwaarden te vergelijken met de bekende digitale stammen. Daarnaast werd de standaardafwijking van de rekwaarden beoordeeld om de heterogeniteit van het rekveld te bepalen. Het verschil tussen de stammen berekend door ALDIC (met behulp van incrementele analyse) en de werkelijke stammen toegepast op de digitaal getransformeerde beelden zijn weergegeven in figuur 1. De gemiddelde spanning in de x-richting berekend door de ALDIC-software was consequent een onderschatting van de werkelijk toegepaste stam (figuur 1A), en de omvang van de fout nam toe met een grotere toegepaste spanning. De magnitude was echter altijd minder dan 0,00015 voor alle rekstappen. Er was ook een lichte onderschatting van de spanning in de y-richting (figuur 1C). De standaardafwijking van de berekende stammen binnen het volledige gebied van belang voor de x-stam en y-stam nam ook toe met grotere toegepaste stammen, maar de magnitude was ook zeer klein (<0,002) (figuur 1B, D). Deze fouten waren aanzienlijk groter bij gebruik van cumulatieve analyse (aanvullende figuur 2).

Figuur 1: Algoritmevergelijking en validatie met digitaal gespannen beelden . (A) De gemeten ALDIC-stamgegevens in de x-richting waren consistent lager dan de werkelijke spanning voorgeschreven door de digitale transformaties, en de fout nam geleidelijk toe met een grotere toegepaste spanning. (B) De standaardafwijking van de rekwaarden in de x-richting nam ook toe bij grotere toegepaste digitale stammen. (C) De gemeten ALDIC-stamgegevens in de y-richting waren consistent lager dan de werkelijke spanning voorgeschreven door de digitale transformaties. (D) De standaardafwijking van de rekwaarden in de y-richting nam toe bij een grotere uitgeoefende spanning. Klik hier om een grotere versie van deze figuur te bekijken.
Bij het uitvoeren van stamanalyse op echte weefselmonsters is het niet mogelijk om de nauwkeurigheid van het ALDIC-algoritme direct te beoordelen. Toch werd een techniek ontwikkeld om de nauwkeurigheid van het verplaatsingsveld in te schatten. In het bijzonder werd het vervormde beeld teruggedraaid tot een voorspelling van het referentiebeeld op basis van het berekende verplaatsingsveld. Een genormaliseerde kruiscorrelatiecoëfficiënt werd vervolgens gebruikt om te bepalen hoe goed het vervormde/gereconstrueerde referentiebeeld overeenkwam met het ware referentiebeeld. Alle subregio's (32 pixels x 32 pixels) waarin de genormaliseerde kruiscorrelatiewaarde minder dan 0,5 was, werden beschouwd als een "slecht gebied" waarin het verplaatsingsveld waarschijnlijk onnauwkeurig was. Uit deze analyse bleek dat er een significant verschil was tussen de prestaties van de incrementele en cumulatieve analysetechnieken. In het bijzonder begon het aantal slechte regio's te stijgen met de cumulatieve methode na de 6% toegepaste stam (figuur 2A), terwijl zeer weinig (1%) slechte regio's werden waargenomen in een van de digitaal getransformeerde regio's voor de incrementele analyse. Bij het toepassen van deze nauwkeurigheidsbeoordelingstechniek op de vier geteste achillespezen van de muis (aanvullende figuur 3), werd vastgesteld dat voor drie monsters het gemiddelde aantal slechte gebieden minder dan 25% van het beeld was. In een van de vier monsters (experiment 2) werd echter bijna de helft van het beeld als slecht geïdentificeerd bij de maximale rektoename (figuur 2B). Het aantal slechte gebieden dat aanwezig was in experiment 2 varieerde van het gemiddelde van de andere drie monsters met meer dan vier standaarddeviaties. Dit maakte het mogelijk om vast te stellen dat de ALDIC-gegevens van experiment 2 een uitschieter vertegenwoordigden, en deze gegevens werden daarom geëlimineerd uit de verdere analyse van de resultaten.

Figuur 2: Succesvolle identificatie van gebieden met ongeldige stamberekeningen door slechte regioanalyse . (A) De hoeveelheid slechte gebieden in de digitaal getransformeerde beelden geanalyseerd met behulp van de cumulatieve methode nam consistent toe na 6% toegepaste spanning, terwijl de incrementele hoeveelheid op 1% bleef. (B) De hoeveelheid slechte gebieden voor alle peesmonsters nam gestaag toe bij grotere rekstappen. Experiment 2 werd beschouwd als een uitschieter en is dus niet opgenomen in de gemiddelde en standaarddeviatiebalken. Klik hier om een grotere versie van deze figuur te bekijken.
Bovendien werden de lokale trekspanningen in de geteste achillespezen van de muis gemeten met behulp van fotogebleekte lijnen (PBL's) als tweede methode voor het bepalen van de nauwkeurigheid van het ALDIC-algoritme. De door ALDIC berekende x-richtingstammen waren meestal groter dan die bepaald uit de PBL's, maar het verschil was over het algemeen binnen 0,005 stam (figuur 3A). Deze foutmagnitude was vergelijkbaar met de standaardafwijking die werd waargenomen tussen de verschillende PBL's binnen een bepaald monster (figuur 3B).

Figuur 3: Validatie van de ALDIC-stamberekeningen door vergelijking met gefotobleekte lijngegevens. (A) Het verschil tussen de ALDIC-stamwaarden en de PBL-stamwaarden bleef relatief constant voor alle rekstijgingen, rond een waarde van 0,005. (B) De standaardafwijking voor de PBL-gegevens, gemiddeld over alle steekproeven, bleef relatief constant op ongeveer 0,005. Klik hier om een grotere versie van deze figuur te bekijken.
Na beoordeling van de nauwkeurigheid van het ALDIC-algoritme werden de groottes en ruimtelijke verdelingen van de lokale stammen in de achillespezen van de muis onder trekbelasting bepaald (figuur 4, figuur 5 en figuur 6). Merk op dat de stammen niet de verplaatsingsgegevens van de "slechte regio's" in elk monster bevatten. De x-richting trekstammen waren consistent over alle drie de monsters en waren aanzienlijk lager dan de toegepaste weefselstammen (figuur 4A). Bovendien was de x-richtingsspanning relatief heterogeen, aangezien de standaarddeviatie over het 2D-beeld altijd groter was dan de gemiddelde rekwaarde. Daarentegen was er een significante inconsistentie tussen de drie monsters voor de y-richtingsstammen, waarbij één monster positieve gemiddelde waarden vertoonde, één monster negatieve gemiddelde waarden vertoonde en één monster nulspanning in de y-richting vertoonde (figuur 4B). Bovendien was de standaarddeviatie van de y-richtingstammen binnen een bepaald monster groter dan de standaardafwijking van de x-richtingstammen. Ten slotte was de schuifspanning relatief laag over alle rekstappen (figuur 4C).

Figuur 4: Microschaalstammen van achillespezen van muizen . (A) De gemiddelde spanning in de x-richting bleef onder de toegepaste weefselstam, maar nam toe met elke toename van de stam. (B) De gemiddelde spanning in de y-richting was ongeveer nul voor alle stappen, maar de standaarddeviatie was hoog. (C) De gemiddelde afschuifspanning nam gestaag toe gedurende de rekstappen. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 5: Ruimtelijke verdeling van x-stammen, y-stammen en afschuifspanningen. Representatieve kaarten van de (A) x-stammen, (B) y-stammen en (C) afschuifspanningen in het hele peesgebied van belang Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 6: Ruimtelijke verdelingen van maximale hoofd-, minimale hoofd- en maximale schuifspanningen. Representatieve kaarten van de (A) maximale hoofdspanningen, (B) minimale hoofdstammen en (C) maximale afschuifspanningen in het hele peesgebied van belang. De witte lijnen geven de richtingen van de maximale en minimale hoofdspanningen aan. Klik hier om een grotere versie van deze figuur te bekijken.
Aanvullende figuur 1: Identificatie van de quasistatische toestand tijdens beeldvorming. De helling van de kracht-tijdcurve tijdens de laatste minuut van de spanningsrelaxatieperiode (rode lijn) kan worden gebruikt om de totale verandering in kracht tijdens de beeldvorming te benaderen. Klik hier om dit bestand te downloaden.
Aanvullende figuur 2: Vergelijking van incrementele en cumulatieve analysetechnieken. (A) Het verschil tussen de gemeten en werkelijke x-richting stammen in de digitaal getransformeerde beelden was significant groter met de cumulatieve methode in vergelijking met de incrementele methode boven 4% spanning. (B) De standaardafwijking van x-stamwaarden was ook significant groter bij de cumulatieve methode boven 4% stam. (C) Het verschil tussen de gemeten en werkelijke y-stammen in de digitaal getransformeerde beelden was aanzienlijk groter met de cumulatieve methode boven 8% spanning. (D) De standaardafwijking van de y-stamwaarden was significant groter bij de cumulatieve methode boven 4% stam. Klik hier om dit bestand te downloaden.
Aanvullende figuur 3: Slechte regiovisualisatie en kwantificering voor elk experiment. Slechte gebieden werden gedefinieerd als lokale gebieden binnen het gereconstrueerde referentiebeeld die niet overeenkwamen (onder de correlatiecoëfficiënt van 0,5) met hetzelfde gebied van het werkelijke referentiebeeld. Elke slechte regio die wordt geïdentificeerd binnen een regio van belang (wit omlijnd) wordt gemarkeerd door een blauw vak. Het percentage slechte regio's binnen de regio van belang wordt boven elke afbeelding tussen haakjes aangegeven. Merk op dat deze afbeeldingen worden gereconstrueerd uit het vervormde beeld bij 12% toegepaste spanning. Klik hier om dit bestand te downloaden.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Het doel van dit artikel was om een open-source, gevalideerde methode te bieden om de 2D-rekvelden in pezen onder trekbelasting te meten. De basis van de software was gebaseerd op een openbaar beschikbaar ALDIC-algoritme12. Dit algoritme was ingebed in een grotere MATLAB-code met de toegevoegde functionaliteit van incrementele (versus cumulatieve) stamanalyse. Dit aangepaste algoritme werd vervolgens toegepast op het trekken van pezen en de nauwkeurigheid ervan werd beoordeeld met twee verschillende technieken (d.w.z. digitaal getransformeerde beelden en rekmeting met behulp van gebleekte lijnen). Bovendien werd een mogelijkheid toegevoegd om de nauwkeurigheid van de ALDIC-metingen op elk monster te beoordelen zonder kennis van de werkelijke stamwaarden te vereisen.
De analyse van de digitaal getransformeerde beelden toonde aan dat het algoritme nauwkeurig spanningen tot 10% kon meten met zeer weinig fouten, die niet konden worden beoordeeld aan de hand van de trekproeven van de werkelijke achillespezen van de muis vanwege de lage omvang van de spanningen in de peesmonsters. Niettemin toonde het vergelijken van de stammen berekend in achillespezen van muizen door ALDIC met de stammen gemeten met behulp van gebleekte lijnen aan dat de fout van de ALDIC-techniek binnen de meetvariatie van de gebleekte lijnen zelf lag. Als laatste validatie werd de nauwkeurigheid van de volledige 2D-verplaatsingsvelden berekend door het ALDIC-algoritme beoordeeld door het referentiebeeld uit het vervormde beeld te reconstrueren en de reconstructie te vergelijken met het werkelijke referentiebeeld. In de digitaal getransformeerde beelden was er een toename van het aantal slechte regio's en rekfouten met grotere toegepaste stammen, vooral voor de cumulatieve ALDIC-analyse (figuur 2 en aanvullende figuur 2). Dit was te verwachten omdat de incrementele techniek het referentiebeeld bij elke tussenliggende afbeelding herdefinieert om de verplaatsingsverschillen tussen afbeeldingsparen te minimaliseren. Het aantal slechte gebieden was nog hoger in de werkelijke peesmonsters omdat de structuur en belasting van het peesweefsel niet homogeen waren (in tegenstelling tot de digitaal getransformeerde beelden). Toch kwam gemiddeld slechts ongeveer 15% van het gereconstrueerde beeld niet overeen met het werkelijke referentiebeeld. Eén steekproef (experiment 2) had echter een groot aantal foutieve regio's (~ 45%). Hoewel het onduidelijk is waarom dit monster niet goed kon worden verwerkt, was deze analyse van het gereconstrueerde referentiebeeld waardevol omdat het de herkenning mogelijk maakte dat de gegevens van dit monster niet betrouwbaar waren. Al met al tonen deze experimenten aan dat dit open-source algoritme met vertrouwen kan worden gebruikt om weefselspanningen in peesexplantaten nauwkeurig te meten.
Deze experimenten leverden ook waardevolle informatie op over het mechanische gedrag van achillespezen van muizen. Specifiek, bij een toegepaste weefselstam van 12%, was de gemiddelde longitudinale (x-richting) stam in het weefselmonster slechts 2%. Een deel van deze rekverzwakking was te wijten aan het feit dat de weefselstammen op macroschaal werden berekend op basis van veranderingen in de grip-to-grip-lengte van het weefsel, die waarschijnlijk significante spanningsconcentraties op het grip-grensvlak van de myotendineuze overgang omvatten. Toch komt dit overeen met andere studies naar microschaalstammen in pezen10,17,18. Bovendien kwam de stam van 12% overeen met ongeveer 5 MPa belasting, wat waarschijnlijk vergelijkbaar is met de maximale fysiologische belasting in vivo19. Dit suggereert dat cellen in de achillespezen van muizen geen trekspanningen van meer dan 2% ervaren. De transversale (y-richting) spanning was meer variabel tussen monsters, met zowel positieve als negatieve waarden. Dit suggereert dat de peesmonsters positieve en negatieve Poisson-verhoudingen vertoonden, wat consistent is met eerdere tests van achillespezen20. Zoals verwacht voor uniaxiale spanning, was de grootte van de schuifspanning over het algemeen laag (gemiddeld <4°). Voor alle trek- en schuifspanningen was de standaarddeviatie in het betreffende gebied echter altijd groter dan de gemiddelde rekwaarde, wat aantoont dat er een grote mate van heterogeniteit van de spanning was. Bovendien nam deze heterogeniteit toe met grotere toegepaste stammen, waarschijnlijk als gevolg van de heterogeniteit van de weefselstructuur en de toegenomen fout binnen de ALDIC-berekeningen als gevolg van de grotere verplaatsings- en verplaatsingsvelden. Dit suggereert dat de stammen die door individuele peescellen worden ervaren, zeer variabel zijn in het weefsel.
Ondanks de succesvolle validatie van het ALDIC-algoritme, zijn er enkele beperkingen in het gebruik ervan voor het analyseren van stammen in peesexplantaten. De belangrijkste beperking is het feit dat het algoritme alleen een 2D-analyse van een 3D-object kan uitvoeren. Een meer rigoureuze aanpak zou zijn om een volledige digitale volumecorrelatie (DVC) uit te voeren, die is uitgevoerd op digitaal getransformeerde beelden van pezen11. Dit is echter over het algemeen moeilijk uit te voeren op echte peesmonsters, omdat de beelden oplosbare kernen bevatten tot een diepte van slechts 100 μm. Dit betekent dat het binnenvolume van de monsters geen textuur heeft binnen de volumetrische beelden, waardoor de DVC onbetrouwbaar is. Daarom werden de beelden in deze studie samengevouwen tot maximale 2D-projecties, die alle kernen kunstmatig in een enkel beeldvlak dwingen. Hoewel dit enkele fouten in de spanningsanalyse kan veroorzaken en de meting van out-of-plane verplaatsingen kan voorkomen, suggereren de validatieresultaten dat de techniek nog steeds nauwkeurig is. Een bijkomende beperking is dat de spanningen werden berekend aan het einde van een spanningsrelaxatieperiode en niet konden worden berekend tijdens dynamische cyclische belasting. Dit probleem was onvermijdelijk omdat er een eindige beeldvormingstijd was om de volumetrische beelden te verkrijgen die voor de stamanalyse werden gebruikt. Ondanks deze beperkingen was het succes van de analyse relatief robuust, aangezien drie van de vier peesmonsters nauwkeurige stamgegevens produceerden. Daarom zal dit algoritme een nuttig hulpmiddel zijn voor onderzoekers die geïnteresseerd zijn in het meten van spanningsvelden binnen peesexplantaten.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Alle auteurs hebben geen belangenconflicten om bekend te maken.
Acknowledgments
Dit werk werd gefinancierd door de National Institutes of Health (R21 AR079095) en de National Science Foundation (2142627).
Materials
| Name | Company | Catalog Number | Comments |
| 5-DTAF (5-(4,6-Dichlorotriazinyl) Aminofluorescein), single isomer | ThermoFisher | D16 | |
| Calipers | Mitutoyo | 500-196-30 | |
| Confocal Microscope | Nikon | A1R HD | |
| Corning LSE Vortex Mixer | Coning | 6775 | |
| DRAQ5 Fluorescent Probe Solution (5 mM) | ThermoFisher | 62554 | |
| MATLAB | MathWorks | R2022b | |
| Tensile Loading Device | N/A | N/A | Tensile loading device described in Peterson et al, 2020. (ref 13) |
| Tube Revolver Rotator | ThermoFisher | 88881001 |
References
- Devkota, A. C. Distributing a fixed amount of cyclic loading to tendon explants over longer periods induces greater cellular and mechanical responses. Journal of Orthopaedic Research. 11 (4), 1609-1612 (2007).
- Sun, H. B., et al. Cycle-dependent matrix remodeling gene expression response in fatigue-loaded rat patellar tendons. Journal of Orthopaedic Research. 28 (10), 1380-1386 (2010).
- Shepherd, J. H., Screen, H. R. C.
- Paschall, L., Pedaprolu, K., Carrozzi, S., Dhawan, A., Szczesny, S. Mechanical stimulation as both the cause and the cure of tendon and ligament injuries. Regenerative Rehabilitation: From Basic Science to the Clinic. , Springer. Cham, Switzerland. 359-386 (2022).
- Andarawis-Puri, N., Ricchetti, E. T., Soslowsky, L. J. Rotator cuff tendon strain correlates with tear propagation. Journal of Biomechanics. 42 (2), 158-163 (2009).
- Cheng, V. W. T., Screen, H. R. C.
- Luyckx, T., et al. Digital image correlation as a tool for three-dimensional strain analysis in human tendon tissue. Journal of Experimental Orthopaedics. 1 (1), 7 (2014).
- Duncan, N. A., Bruehlmann, S. B., Hunter, C. J., Shao, X., Kelly, E. J. In situ cell-matrix mechanics in tendon fascicles and seeded collagen gels: Implications for the multiscale design of biomaterials. Computer Methods in Biomechanics and Biomedical Engineering. 17 (1), 39-47 (2014).
- Arnoczky, S. P., Lavagnino, M., Whallon, J. H., Hoonjan, A. In situ cell nucleus deformation in tendons under tensile load; A morphological analysis using confocal laser microscopy. Journal of Orthopaedic Research. 20 (1), 29-35 (2002).
- Screen, H. R. C., Bader, D. L., Lee, D. A., Shelton, J. C.
- Fung, A. K., Paredes, J. J., Andarawis-Puri, N. Novel image analysis methods for quantification of in situ 3-D tendon cell and matrix strain. Journal of Biomechanics. 67, 184-189 (2018).
- Yang, J., Bhattacharya, K. Augmented Lagrangian digital image correlation. Experimental Mechanics. 59 (2), 187-205 (2019).
- Peterson, B. E., Szczesny, S. E. Dependence of tendon multiscale mechanics on sample gauge length is consistent with discontinuous collagen fibrils. Acta Biomaterialia. 117, 302-309 (2020).
- Humphrey, J. D., O'Rourke, S. L. An Introduction to Biomechanics. , Springer. New York, NY. (2015).
- Reese, S. P., Weiss, J. A. Tendon fascicles exhibit a linear correlation between Poisson's ratio and force during uniaxial stress relaxation. Journal of Biomechanical Engineering. 135 (3), 34501 (2013).
- Ahmadzadeh, H., Freedman, B. R., Connizzo, B. K., Soslowsky, L. J., Shenoy, V. B. Micromechanical poroelastic finite element and shear-lag models of tendon predict large strain dependent Poisson's ratios and fluid expulsion under tensile loading. Acta Biomaterialia. 22, 83-91 (2015).
- Szczesny, S. E., Elliott, D. M. Interfibrillar shear stress is the loading mechanism of collagen fibrils in tendon. Acta Biomaterialia. 10 (6), 2582-2590 (2014).
- Han, W. M., et al. Macro- to microscale strain transfer in fibrous tissues is heterogeneous and tissue-specific. Biophysical Journal. 105 (3), 807-817 (2013).
- Pedaprolu, K., Szczesny, S. E. A novel, open-source, low-cost bioreactor for load-controlled cyclic loading of tendon explants. Journal of Biomechanical Engineering. 144 (8), 084505 (2022).
- Gatt, R., et al. Negative Poisson's ratios in tendons: An unexpected mechanical response. Acta Biomaterialia. 24, 201-208 (2015).
