Vector Competence Analyses on Aedes aegypti Mosquitoes using Zika Virus
Summary
The presented protocol can determine the vector competence of Aedes aegypti mosquito populations for a given virus, such as Zika, in a containment setting.
Abstract
The procedures presented describe a generalized methodology to infect Aedes aegypti mosquitoes with Zika virus under laboratory conditions to determine the rate of infection, disseminated infection, and potential transmission of the virus in the mosquito population in question. These procedures are widely utilized with various modifications in vector competence evaluations globally. They are important in determining the potential role that a given mosquito (i.e., species, population, individual) may play in the transmission of a given agent.
Introduction
Vector competence is defined as the ability at the level of species, population, and even an individual, of a given arthropod such as a mosquito, tick, or phlebotomine sand fly, to acquire and transmit an agent biologically with replication or development in the arthropod1,2. With respect to mosquitoes and arthropod-borne viruses (i.e., arboviruses), the agent is imbibed from a viremic host by a female mosquito. Following ingestion, the virus must productively infect one of a small population of midgut epithelial cells3, overcoming various physiological obstacles such as proteolytic degradation by digestive enzymes, the presence of the microbiota (midgut infection barrier, or MIB), and the secreted peritrophic matrix. Infection of the midgut epithelium must be followed by replication of the virus and eventual escape from the midgut into the mosquito’s open circulatory system, or hemolymph, which represents the onset of a disseminated infection overcoming the midgut escape barrier (MEB). At this point the virus can establish infections of secondary tissues (e.g., nerves, muscles, and fat bodies) and continue to replicate, although such secondary replication may not be strictly necessary for the virus to infect the acinar cells of the salivary glands (overcoming the salivary gland infection barrier). Egress from the salivary gland acinar cells into their apical cavities and then movement into the salivary duct enables inoculation of the virus into subsequent hosts on biting, and completes the transmission cycle1,2,4,5,6,7.
Given this well-characterized and generally conserved mechanism of spread within a mosquito vector, laboratory vector competence assessments are often methodologically similar, although differences in protocols do exist1,2. Generally, after oral virus exposure, mosquitoes are dissected so that individual tissues such as the midgut, legs, ovaries, or salivary glands can be assayed for viral infection, disseminated infection, disseminated infection/potential transovarial transmission, and disseminated infection/potential transmission competence, respectively8. The mere presence of a virus in the salivary glands, however, is not definitive evidence of transmission capability, given evidence of a salivary gland escape/egress barrier (SGEB) in some vector/virus combinations1,2,4,5,7,9. The standard method to prove transmission competence remains mosquito transmission to a susceptible animal10,11,12. However, given that for many arboviruses this necessitates the use of immunocompromised murine models13,14,15,16, this method is often cost-prohibitive. A commonly used alternative is the collection of the mosquito saliva, which can be analyzed by reverse transcription-polymerase chain reaction (RT-PCR) or an infectious assay to demonstrate the presence of the viral genome or infectious particles, respectively. It is worth noting that such in vitro saliva collection methods may overestimate12 or underestimate17 the amount of virus deposited during in vivo feeding, indicating that such data must be interpreted with caution. Nonetheless, the in vitro method is highly valuable when analyzed from the perspective of the mere presence of virus in the saliva, indicating transmission potential.
Two major approaches exist for determining the role of mosquito vectors in arboviral disease outbreaks. The first method involves field surveillance, in which mosquitoes are collected in the context of active transmission18,19,20,21,22,23,24. However, given that infection rates are typically quite low (e.g., the estimated 0.061% infection rate of mosquitoes in areas of active Zika virus (ZIKV) circulation in the United States21), incrimination of potential vector species can be heavily biased by trapping methodology25,26 and random chance (e.g., sampling one infected individual out of 1,600 uninfected)21. Taking this into account, a given study may not acquire sufficient mosquitoes in both raw numbers or species diversity to accurately sample mosquitoes involved in transmission. In contrast, vector competence analyses are undertaken in a laboratory setting, allowing for strict control of parameters such as oral dose. Although not fully capable of representing the true complexity of mosquito infection and transmission capability in a field setting, these laboratory assessments remain powerful tools in the field of arbovirology.
Based on various vector competence analyses with ZIKV in several mosquito species, populations, and methods27,28,29,30,31,32, as well as a recent review of vector competence assessments1, we describe here several of the protocols associated with a typical vector competence workflow. In these experiments, three Ae. aegypti populations from the Americas (the city of Salvador, Brazil; the Dominican Republic; and the lower Rio Grande Valley, TX, USA) were exposed to a single strain of ZIKV (Mex 1-7, GenBank Accession: KX247632.1) at 4, 5, or 6 log10 focus-forming units (FFU)/mL doses by way of artificial bloodmeals. Subsequently, they were analyzed for evidence of infection, disseminated infection, and transmission competence after various times of extrinsic incubation (2, 4, 7, 10, and 14 days) by means of dissection and a cell culture-based infectious assay. Although the present workflow/protocols are optimized for ZIKV, many elements are directly translatable to other mosquito-borne arboviruses in arthropod containment and biosafety levels 2 and 3 (ACL/BSL2 or ACL/BSL3).
Protocol
All procedures performed in these protocols were performed in full compliance with protocols approved by the Institutional Biosafety Committee and the Institutional Animal Care and Use Committee of the University of Texas Medical Branch at Galveston.
1. Amplify ZIKV in Vero cells
- Grow Vero cells (CCL-81 or VeroE6) in Dulbecco’s modification of Eagle’s minimal essential medium (DMEM) supplemented with 10% v/v heat-inactivated fetal bovine serum (FBS), and 1% (v/v) penicillin-streptomycin (100 U/mL and 100 μg/mL, respectively) in a humidified 37 °C incubator with 5% CO2 to between 80−90% confluency in a 150 cm2 tissue culture flask.
- In a biosafety cabinet (BSC), remove the medium and dispose of in either 10% bleach or a working dilution of dual quaternary ammonium (Table of Materials). Immediately inoculate the monolayer with 1 mL of viral stock, aiming for 0.1−1 infectious viral particle per cell. Agitate the flask immediately such that the inoculum contacts the monolayer in its entirety.
- Top up the medium to a volume of 5 mL using DMEM supplemented with 2% v/v heat-inactivated FBS, and 1% (v/v) penicillin-streptomycin. Then move the flask into the humidified 37 °C incubator with 5% CO2 for 60 min.
- Remove the flask from the incubator and bring it into a BSC. Add additional medium to a total volume of 15 mL using DMEM supplemented with 2% v/v heat-inactivated FBS, and 1% (v/v) penicillin-streptomycin. Move the flask into a humidified 37 °C incubator with 5% CO2.
- Examine the flask daily under phase contrast microscopy for evidence of cytopathic effects (CPE). Proceed to the viral harvest (step 1.7) when only approximately 40−50% of the cells remain in the monolayer, generally 3−5 days postinfection depending on the strain of ZIKV being utilized.
- Aspirate the supernatant and place in a 50 mL conical vial. Clarify the supernatant of cellular debris by centrifugation (3,500 x g for 20 min).
NOTE: If multiple flasks were infected identically, supernatants from multiple flasks can be combined into 50 mL conical tubes. - Remove the supernatant from the 50 mL conical tube to a fresh one, taking care not to disrupt the pellet. Supplement the supernatant with heat-inactivated FBS to a final concentration of 30% (v/v). Aliquot this mixture into individual screw cap tubes and freeze at -80 °C until use.
2. Preparation of artificial bloodmeals
- On the day mosquitoes are to be exposed to infectious bloodmeals, turn on the power source (Table of Materials) in an arthropod containment facility such that the feeding units (Table of Materials) are preheated by the time the mosquitoes are prepared for exposure.
- Prepare artificial bloodmeals using one of the methods described below.
- Method 1: Combine freshly harvested (within a week) citrated or heparinized human blood purchased commercially 1:1 v/v with viral stock (prepared as described in section 1).
NOTE: This method is contingent on the absence of any antibodies to the virus/virus family being present in the blood. - If absence of antibodies cannot be confirmed or the blood source is known to have prior flavivirus exposure, wash and manually pack erythrocytes.
- In a BSC, add 30 mL of whole human blood to a 50 mL conical tube and top up to 50 mL with phosphate buffered saline (PBS).
- Centrifuge at 3,500 x g for 20 min.
- Aspirate the supernatant either by gently pouring into a tray pan containing either 10% bleach or working dilution of dual quaternary ammonium, taking care to not discard the erythrocyte pellet.
- Add 10 mL of PBS and gently tap the bottom of the conical tube against the bottom of the BSC such that the erythrocyte pellet has been reconstituted. Bring the volume of the suspension up to 50 mL with PBS. Mix by gentle inversion.
- Repeat steps 2.2.2.1−2.2.2.4 a total of 4−6x. Confirm that the supernatant is clear or only slightly pink and no longer opaque. Remove all the supernatant using a serological pipette. Resuspend the erythrocyte pellet in 1−2 mL of PBS.
- Assemble the bloodmeal: 350 μL of packed erythrocytes, 100 μL of 10% sucrose, 200 μL of heat-inactivated FBS, 900 μM recombinant ATP, and 2 mL of appropriately diluted virus stock using DMEM supplemented with 2% v/v heat-inactivated FBS, and 1% (v/v) penicillin-streptomycin.
- Method 1: Combine freshly harvested (within a week) citrated or heparinized human blood purchased commercially 1:1 v/v with viral stock (prepared as described in section 1).
- Overlay a standard 3 mL reservoir unit (Table of Materials) with the skin of an uninfected mouse (other options include paraffin film, collagenous membranes, or sausage casing).
- Place the covered reservoir on white paper towels. Add ~2 mL of infectious bloodmeal to the reservoir 1 mL at a time. Inspect the towel underneath the feeder for any evidence of leakage. If leaks are present, recover the bloodmeal from the feeders and discard the covers. Seal the feeders with plugs. Once again confirm that no leaks are present.
3. Backtitration of bloodmeals/plaque assay
- Using the remaining volume of the prepared bloodmeal, perform a 10x serial dilution series (i.e., 6 dilutions, ranging from diluted 10x to 1,000,000x) using DMEM supplemented with 2% v/v heat-inactivated FBS, and 1% (v/v) penicillin-streptomycin.
- Aliquot 100 µL of the dilutions into the wells of 24 or 12 well plates from the most dilute to most concentrated.
- Incubate for 1 h in a 37 °C, 5% CO2 incubator.
- At the end of the 1 h incubation period, bring the plates back into the BSC and add 1 mL or 2 mL of methylcellulose overlay to the 24 well or 12 well plates, correspondingly.
- Place overlayed plates back in a 37 °C, 5% CO2 incubator and incubate for 3−7 days (virus strain-dependent).
- Following incubation, remove the plates from the incubator and bring into the BSC. Discard the methylcellulose overlay into a tray pan containing 10% bleach or dual quaternary ammonium disinfectant.
- Wash each well 2x with PBS, discarding the wash into a tray pan containing 10% bleach or dual quaternary ammonium.
- Add ~1 mL of methanol:acetone (1:1 v:v) and allow the cells to fix onto plate for at least 30 min in the BSC at room temperature (RT). Discard methanol:acetone according to institutional policy on organic waste.
- Visualize ZIKV using one of two methods described below.
- Following the removal of methanol:acetone, stain immediately with a crystal violet solution (0.25% w/v in 30% methanol) for 5 min. Rinse 2x in tap water and leave to dry, then directly visualize by eye for evidence of plaques or destruction of monolayer.
- Alternatively, perform focus forming assay.
- Allow the plates to air dry until no organic fixative remains.
NOTE: This should take ~2−3 h outside of the BSC but can be accelerated by air-drying in a BSC or chemical fume hood. - Wash each well 3x for 15 min each in nonsterile PBS (Mg2+ and Ca2+ free) on an orbital plate rocker. Remove PBS and add 1 mL of blocking solution (PBS + 3% FBS) to each well and rock for 15 min at RT.
- Add 100 μL per well of α-ZIKV or α-flavivirus primary antibody (e.g., flavivirus group hybridoma D1-4G2-4-15 [4G2]) at a 1:2,000 dilution in blocking solution. Incubate with rocking for a minimum of 4 h (preferably overnight, not exceeding 18 h) at RT.
- Remove the primary antibody and wash 3x for 15 min each with PBS (Mg2+ and Ca2+ free) on an orbital plate rocker.
- Add 100 µL per well of the secondary antibody (goat α mouse HRP-labeled) diluted 1:2,000 in blocking buffer. Incubate with rocking for 1 h at RT.
- Wash 3x for 15 min each with PBS (Mg2+ and Ca2+ free) on an orbital plate rocker.
- Aliquot 100 µL of substrate development reagent (Table of Materials) per well. Rock plates at RT for 15 min.
- Halt the reaction upon development of foci/plaques by removing the substrate and rinsing the plates 2x with tap water. Pour off tap water and allow the plates to air dry before quantifying.
- Count viral foci to determine the number of FFU present in the given sample.
- Allow the plates to air dry until no organic fixative remains.
4. Administration of bloodmeals
- Use Ae. aegypti mosquitoes 2−4 days post eclosion. Sort female mosquitoes into 0.5 L cardboard cartons with screened lids and deprive them of sugar (generally 36−48 h prior to the infectious bloodmeal). Provide ad libitum water via water-saturated cotton balls.
- Remove the water-saturated cotton balls on the morning that the mosquitoes will be exposed to the infectious bloodmeal.
- Attach reservoirs containing artificial bloodmeals to feeding unit leads within a clear plastic glove box.
- Within the glovebox, place a 0.5 L cardboard carton with a screened lid, containing 50−100 starved Ae. aegypti mosquitoes, underneath the feeding unit attached to the reservoir.
NOTE: Appropriately starved mosquitoes will generally feed within 20 min. Feeding can be prolonged as needed to increase sample size in slower populations, although this should not extend beyond 60 min, because (ZIKV) viral titer can decrease within the feeder after ~60 min. - Upon completion of feeding, remove the reservoir and immerse in freshly made 10% bleach.
- Cold-anesthetize mosquitoes by incubation for 30 s at -20 °C or for 5 min in a refrigerator.
- Within the glovebox, pour the mosquitoes into a Petri dish on ice. Count and sort the engorged females from unengorged mosquitoes. Dispose of the unengorged mosquitoes by immersion in a 50 mL tube conical tube filled with 70% ethanol. While the mosquitoes are still anesthetized, pour them back into the 0.5 L cardboard carton and quickly cover with the screen and lid. Trim excess screen mesh off of the carton and secure the mesh with tape.
- Add a cotton ball saturated with sterile filtered 10% aqueous sucrose to the screen of each carton. Place all mosquito cartons in a large plastic secondary container with a damp sponge to maintain humidity.
- Place secondary container containing cartons of mosquitoes into an incubator with a temperature of 27 ± 1 °C (or as appropriate to simulate conditions in region of interest) with 80% ± 10% relative humidity and a 16:8 light:dark cycle). Maintain the mosquitoes with ad libitum access to 10% sucrose until completion of the experiments.
5. Sample acquisition and processing
- On specified days postfeeding, aspirate a predetermined number of mosquitoes from the appropriate cartons using a mechanical aspirator within a glovebox. Cap the collection tube with a cotton round after the requisite number of mosquitoes is acquired.
- Cold-anesthetize mosquitoes by incubation for 30 seconds at -20 °C or for 5 minutes at 4 °C.
- Within the glovebox, pour the mosquitoes into a Petri dish on ice. Using two pairs of forceps, remove all six of each mosquito’s legs and place in a prelabeled 2 mL round bottom microcentrifuge tube containing a sterilized stainless steel ball bearing (7/32”) and 500 μL of mosquito collection media (MCM) composed of DMEM, 2% FBS, 1% penicillin-streptomycin, and 2.5 μg/mL amphotericin.
- Gently place the mosquito onto a drop of mineral oil to restrain it, taking care to not allow any contact between the oil and the mosquito’s head and proboscis.
- Insert the mosquito proboscis into a 10 μL pipette tip filled with 10 μL of heat-inactivated FBS.
NOTE: Alternatively, the pipette can be filled with sucrose, blood, or mineral oil. The oil allows for direct visualization of saliva bubbles via light microscopy. - Allow the mosquito to salivate for 30 min. Eject the micropipette tip containing FBS + saliva into a microcentrifuge tube containing 100 μL of MCM, then place the carcass into a separate 2 mL round bottom microcentrifuge tube containing a sterilized steel ball bearing and 500 μL of MCM. Ensure that the tubes used for the bodies, legs, and saliva are labeled so that it is clear all three samples originated from the same mosquito.
- While the mosquito(s) are salivating, perform steps 5.3−5.5 on the remaining mosquitoes.
- Transport the tubes containing the bodies and legs to a bead milling tissue homogenization device contained within a BSC. Triturate all body and leg samples at 26 Hz for 5 min to liberate viral particles into the supernatant. Clarify all samples by centrifugation at 200 x g for 5 min to pellet cellular debris.
NOTE: At this point, samples can be frozen at -80 °C, or assayed immediately.
6. Detection of ZIKV by infectious assay
- In a BSC, prepare 24 well tissue culture plates with Vero cells (105 cells per well) 24 h prior to the onset of the infectious assay. Label each well with the identity of a single mosquito/sample.
- If samples were frozen at -80 °C, allow to thaw.
- Remove the media from the Vero cell plates prior to inoculation with samples one plate at a time.
- For samples containing bodies or legs, carefully aliquot 100 μL of clarified supernatant into each well, taking care not to disturb the mosquito cell debris from the pellet.
NOTE: Saliva samples can be diluted 1:1 (v/v) with MCM prior to inoculation onto cells to conserve samples for later titration, if necessary. - Move the plates into a 37 °C, 5% CO2 incubator and incubate for 1 h.
- Return the plates to the BSC and add ~1 mL of methylcellulose overlay to each well. Place the overlayed plates back in a 37 °C, 5% CO2 incubator and incubate for 3−7 days (virus/strain-dependent).
- Perform fixation and visualization as described in steps 3.6−3.9.
- Regarding scoring via a focus forming assay, quantify the positive wells by examination under a light microscope. Detection of intracytoplasmic staining of cells in a well indicates the presence of virus. Samples are scored solely as focus-positive or -negative.
Representative Results
Three populations of Ae. aegypti from the Americas (Salvador, Brazil; the Dominican Republic; and the Rio Grande Valley, TX, USA) were exposed to an outbreak strain of ZIKV from the Americas (ZIKV Mex 1-7, Chiapas State, Mexico, 2015) over a range of bloodmeal titers (4, 5, and 6 log10 FFU/mL) presented in a washed human erythrocyte-based artificial bloodmeal. At days 2, 4, 7, 10, and 14 postinfection, subsets of mosquitoes were processed to determine infection, dissemination, and potential transmission rates.
At a bloodmeal titer of 4 log10 FFU/mL of ZIKV Mex 1−7, Ae. aegypti from Salvador, Brazil were infected at rates of 12.5% and 11.1% after 4 and 14 days of extrinsic incubation, respectively, with no evidence of disseminated infection observed in the assayed legs, and no virus detected in the saliva (Figure 1a). Increasing the titer to 5 log10 FFU/mL resulted in a marginal increase in infectivity with rates of 22.2%, 33.3%, and 22.2% on days 4, 10, and 14 postinfection, respectively. Similar to what was observed in the cohort exposed to 4 log10 FFU/mL, no disseminated infections or transmission competence was observed at any time point (Figure 1b). At the highest examined titer (6 log10 FFU/mL) no infections were identified after 2 days of incubation, but infections were observed at all other time points, peaking at 88.9% by 10 days of extrinsic incubation. ZIKV was detected in the legs of mosquitoes examined at 10 and 14 days postinfection (22.2% and 66.7%, respectively), indicating that ZIKV had disseminated into the hemocoel, although infectious ZIKV was observed in saliva at these time points (Figure 1c).
The Ae. aegypti population from the Dominican Republic proved the most susceptible to ZIKV infection and were transmission competent after exposure to all tested bloodmeal titers. Some level of infection was observed at all time points at all three tested doses, with the lowest rate observed 2 days postinfection at 4 log10 FFU/mL (25%) (Figure 1d). With bloodmeal titers of 5 log10 and 6 log10 FFU/mL conditions infection rates peaked at 100%, with 100% infection observed as early as 4 days postinfection in the population of mosquitoes exposed to 6 log10 FFU/mL ZIKV (Figure 1e,f). Mosquitoes fed all three doses demonstrated dissemination by 7 days postinfection, peaking at 44.4% (4 log10 FFU/mL, 14 days postinfection), 88.9% (5 log10 FFU/mL, 1 and 14 days postinfection), and 100% (6 log10 FFU/mL, 10 days postinfection). Transmission-competence was observed after all three doses (11.1%, 22.2%, and 22.2% at 4, 5, and 6 log10 FFU/mL respectively), but only following a 14 day extrinsic incubation period (EIP) (Figure 1d−f).
The Ae. aegypti population from the Rio Grande Valley, TX, proved to be relatively refractory to infection with ZIKV. Mosquitoes exposed to bloodmeal titers of 4 log10 FFU/mL, were infected as early as 4 days postinfection, with infection rates between 22.2% and 44.4%. With these exposure conditions, disseminated infections were observed at 14 days postinfection at a rate of 11.1%, and no transmission competence was observed (Figure 1g). A ten-fold increase in bloodmeal titer produced a largely similar result, with infections observed starting 4 days postexposure (33.3% and 44.4%), while disseminated infections were found after 14 days of extrinsic incubation at a rate of 22.2% (Figure 1h). Finally, in the cohort exposed to a 6 log10 FFU/mL bloodmeal, infection was observed beginning from the 2 day postinfection time point (22.2%) and reached peaks at 4, 10, and 14 days postinfection (66.7%). Disseminated infections in this condition began to be observed at 7 days postinfection (11.1%) and peaked at 44.4% at 14 days postinfection. Only a single mosquito (11.1%) was observed to be transmission capable at 10 days postinfection (Figure 1i).
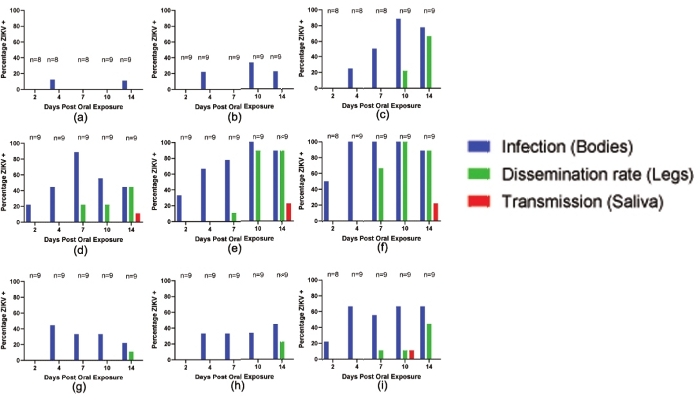
Figure 1: Representative vector competence data of various Ae. aegypti populations for ZIKV Mex 1−7. (a−c) Vector competence of Ae. aegypti from Salvador, Brazil (F2). (d−f) Vector competence of Ae. aegypti from the Dominican Republic (F6). (g−i) Vector competence of Ae. aegypti from the Rio Grande Valley, TX (F4). (a,d,g) Ae. aegypti exposed to 4 log10 FFU/mL of ZIKV Mex 1-7. (b,e,h) Ae. aegypti exposed to 5 log10 FFU/mL of ZIKV Mex 1-7. (c,f,i) Ae. aegypti exposed to 6 log10 FFU/mL of ZIKV Mex 1-7. At each time point (2, 4, 7, 10, and 14 days postinfection) a subset of mosquitos was collected and sampled. Infection, dissemination, and transmission rates are presented as the number of positive carcass/leg/saliva samples over the number of mosquitos assayed at that time point. Infection represented in blue, disseminated infections represented in green, and transmission rate represented in red. Data in this figure are modified from Roundy and Azar et al.32. Please click here to view a larger version of this figure.
Discussion
The methods described here provide a generalized workflow to conduct vector competence analyses. As a general framework, many of these methodologies are conserved throughout the literature. However, there is substantial room for modifications (reviewed in Azar and Weaver1). Virus (e.g., viral lineage, storage of challenge virus, viral passage history), entomology (e.g., laboratory colonization of mosquito populations, innate immunity, the mosquito microbiome/virome), and experimental variables (e.g., bloodmeal composition, sequential blood feeding, and incubation temperature) are all known to affect vector competence. Methodological variability in competence studies has proven problematic in the context of the ZIKV outbreak because it has precluded formal meta-analyses1,33.
Within this general methodology, the importance of the appropriate starvation of mosquitoes and bloodmeal makeup cannot be overstated. Dehydration is known to drive blood feeding behavior of mosquitos in laboratory paradigms34, underscoring the value of sugar and water starvation prior to offering an infectious bloodmeal. While deprivation of sugar for 36–48 h and water for 2–4 h prior to exposure to the bloodmeal is well tolerated by Ae. aegypti, it is worth noting that these mosquitoes are notoriously easy to work with in laboratory conditions2. Such aggressive regimens of starvation may not be nearly as well tolerated by other mosquito species, necessitating some degree of in-house optimization. Likewise, bloodmeal contents may be informed by host preference. For example, using human blood to prepare a bloodmeal for anthropophilic mosquitos like Ae. aegypti is entirely appropriate, but human bloodmeals made for ornithophilic species such as Culex quinquefasciatus may prove less effective35. Additionally, with respect to bloodmeal assembly, the use of relatively fresh blood products is highly advisable to minimize hemolysis of erythrocytes.
One of the greatest limitations of vector competence assessment as a whole and the procedures described herein is that these studies are largely limited to investigating viruses using mosquitoes that can be maintained in laboratory conditions. Ae. aegypti, while a highly relevant vector for a multitude of pathogens of clinical importance, also happens to be one of the easiest mosquitoes to rear and maintain in laboratory colonies1,31,36,37. Unsurprisingly, the competence of Ae. aegypti populations is often therefore the best characterized among common vector mosquitoes2. This is particularly problematic in the context of arboviruses that maintain both enzootic and urban transmission cycles38,39,40, as vector competence is generally only conducted in the context of the more tractable urban mosquitoes.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
We acknowledge the staff of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA): Dr. Robert Tesh, Hilda Guzman, Dr. Kenneth Plante, Dr. Jessica Plante, Dionna Scharton, and Divya Mirchandani, for their tireless work in curating and providing many of the viral strains used for our and other groups’ vector competence experiments. The presented work was funded by the McLaughlin Fellowship Fund (SRA) and NIH grants AI120942 and AI121452.
Materials
| 3mL Standard Reservoir | R37P30 | Hemotek Ltd | Insectary Equipment |
| 7/32" Stainless Steel 440 Grade C Balls | 4RJH9 | Grainger | Grinding Media |
| Acetone, Histological Grade, Fisher Chemicals, Poly Bottle, 4L, 4/Case | A16-P4 | FisherScientific | Fixative |
| Adenosine 5'-triphospate disodium salt hydrat, microbial, BioReagent, suitable for cell culture | A6419-1G | MilliporeSigma | Reagent |
| Anti-Flavivirus Group Antigen Antibody, clone D1-4G2-4-15 | MAB10216 | MilliporeSigma | Primary Antibody for focus forming assay |
| Anti-Mouse IgG (H+L) Antibody, Human Serum Adsorbed and Peroxidase-Labeled, 1.0mL/Bottle | 5450-0011 | KPL/Seracare | Secondary Antibody for focus forming assay |
| Bleach | NC0427256 | FisherScientific | Decontamination |
| Corning, Cell Culture Treated Flasks, 150cm2, Vented Cap, Case of 50 | 10-126-34 | FisherScientific | Cell culture consumable |
| Costar Cell Culture Plates, 24-well, 5/bag, 100/case, Corning | 07-200-740 | FisherScientific | Cell culture consumable |
| Costar Cell Culture Plates, 96-well, 5/bag, 100/case, Corning | 07-200-91 | FisherScientific | Cell culture consumable |
| Crystal Violet | C0775-100G | MilliporeSigma | Stain |
| Eppendorf Snap Cap Microcentrifuge Safe-Lock 2mL Tubes, 500/Case | 05-402-7 | FisherScientific | Plastic consumable |
| Falcon 15mL Conical Centrigue Tubes | 14-959-70C | FisherScientific | Plastic consumable |
| Falcon 50mL Conical Centrigue Tubes | 14-959-49A | FisherScientific | Plastic consumable |
| Falcon Disposable Polystyrene Serological 10mL Pipets, 200/Case | 13-675-20 | FisherScientific | Plastic consumable |
| Falcon Disposable Polystyrene Serological 1mL Pipets, 1000/Case | 13-675-15B | FisherScientific | Plastic consumable |
| Falcon Disposable Polystyrene Serological 25mL Pipets, 200/Case | 13-675-30 | FisherScientific | Plastic consumable |
| Falcon Disposable Polystyrene Serological 5mL Pipets, 200/Case | 13-675-22 | FisherScientific | Plastic consumable |
| Falcon Standard Tissue Culture Dishes | 08-772B | FisherScientific | Plastic consumable |
| Fetal Bovine Serum-Premium, 500mL | S11150 | Atlanta Biologicals | Cell culture reagent |
| Fisherbrand Economy Plain Glass Microscope Slides | 12-550-A3 | FisherScientific | Immobilization of Mosquitos |
| FU1 Feeder | FU1-0 | Hemotek Ltd | Insectary Equipment; feeding units |
| Gibco DPBS with Calcium and Magnesium, 10 x 500mL Bottles | 140-040-182 | FisherScientific | Cell culture reagent |
| Gibco Fungizone, Amphotericin B, 250μg/mL, 50mL/Bottle | 15-290-026 | Fisher Scientific | Cell culture reagent |
| Gibco Penicillin-Streptomycin (10,000 U/mL), 100mL/Bottle, 20 Bottles/Case | 15-140-163 | FisherScientific | Cell culture reagent |
| Gibco, Tryptsin-EDTA (.25%), Phenol red, 20 x 100mL Bottles | 25-200-114 | FisherScientific | Cell culture reagent |
| Gibcom DMEM, High Glucose, 10 x 500mL Bottles | 11-965-118 | FisherScientific | Cell culture reagent |
| Human Blood, Unspecified Gender, Na-Citrate, 1 Unit | 7203706 | Lampire | Bloodmeal preparation |
| InsectaVac Aspirator | 2809B | Bioquip | Insectary Equipment |
| Methanol, Certified ACS, Fisher Chemicals, Amber Glass Bottle, 4L, 4/Case | A412-4 | FisherScientific | Fixative |
| Methyl cellulose, viscosity: 3,500-5,600 cP, 2 % in water(20 °C), 250g/Bottle | M0512-250G | MilliporeSigma | Cell culture reagent |
| Micro-chem Plus Disinfectant Detergent | C849T34 | Thomas Scientific | Decontamination; working dilution of dual quaternary ammonium |
| Mineral Oil, BioReagent, for molecular biology | M5904-5X5ML | MilliporeSigma | Immobilization of Mosquitos |
| O-rings | OR37-25 | Hemotek Ltd | Insectary Equipment |
| Plastic Plugs | PP5-250 | Hemotek Ltd | Insectary Equipment |
| PS6 Power Unit (110-120V) | PS6120 | Hemotek Ltd | Insectary Equipment; power source |
| Rubis Forceps, Offset blades, superfine points | 4525 | Bioquip | Insectary Equipment |
| Sarstedt Inc, 2mL Screw Cap Microtube, Conical Bottom, O-ring Cap, Sterile, 1000/Case | 50-809-242 | FisherScientific | Plastic consumable |
| Sucrose, BioUltra, for molecular biology | 84097-250G | MilliporeSigma | Reagent |
| ThermoScientific, ART Barrier Low Retention 1000μL Pipette Tips, 100 tips/Rack, 8 Racks/Pack, 4 Packs/Case | 21-402-487 | FisherScientific | Plastic consumable |
| ThermoScientific, ART Barrier Low Retention 200μL Pipette Tips, 96 tips/Rack, 10 Racks/Pack, 5 Packs/Case | 21-402-486 | FisherScientific | Plastic consumable |
| ThermoScientific, ART Barrier Low Retention 20μL Pipette Tips, 96 tips/Rack, 10 Racks/Pack, 5 Packs/Case | 21-402-484 | FisherScientific | Plastic consumable |
| ThermoScientific, ART Barrier Low Retention, Extended Reach 10μL Pipette Tips, 96 tips/Rack, 10 Racks/Pack, 5 Packs/Case | 21-402-482 | FisherScientific | Plastic consumable |
| TissueLyser II | 85300 | QIAGEN | Homogenization |
| TrueBlue Peroxidase Substrate Kit, 200mL | 5510-0030 | Seracare | Developing solution for focus forming assay |
| Vero | CCL-81 | American Type Culture Collection | Mammalian cell line to amplify virus and conduct infectious assay |
| Vero C1008 [Vero 76, clone E6, Vero E6] | CRL-1586 | American Type Culture Collection | Mammalian cell line to amplify virus and conduct infectious assay |
Riferimenti
- Azar, S. R., Weaver, S. C. Vector Competence: What Has Zika Virus Taught Us. Viruses. 11 (9), 867 (2019).
- Souza-Neto, J. A., Powell, J. R., Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infection, Genetics and Evolution. 67, 191-209 (2019).
- Smith, D. R., Adams, A. P., Kenney, J. L., Wang, E., Weaver, S. C. Venezuelan Equine Encephalitis Virus in the Mosquito Vector Aedes taeniorhynchus: Infection Initiated by a Small Number of Susceptible Epithelial Cells and a Population Bottleneck. Virology. 372 (1), 176-186 (2008).
- Forrester, N. L., Coffey, L. L., Weaver, S. C. Arboviral bottlenecks and challenges to maintaining diversity and fitness during mosquito transmission. Viruses. 6 (10), 3991-4004 (2014).
- Kramer, L. D., Ciota, A. T. Dissecting vectorial capacity for mosquito-borne viruses. Current Opinion in Virology. 15, 112-118 (2015).
- Kramer, L. D., Hardy, J. L., Presser, S. B., Houk, E. J. Dissemination Barriers for Western Equine Encephalomyelitis Virus in Culex tarsalis infected after Ingestion of Low Viral Doses. American Journal of Tropical Medicine and Hygiene. 30 (1), 190-197 (1981).
- Lounibos, L. P., Kramer, L. D. Invasiveness of Aedes aegypti and Aedes albopictus and Vectorial Capacity for Chikungunya Virus. The Journal of Infectious Diseases. 214, 453-458 (2016).
- Heitmann, A., et al. Forced Salivation as a Method to Analyze Vector Competence of Mosquitoes. Journal of Visualized Experiments. (138), e57980 (2018).
- Beerntsen, B. T., James, A. A., Christensen, B. M. Genetics of Mosquito Vector Competence. Microbiology and Molecular Biology Reviews. 64 (1), 115-137 (2000).
- Guo, X. X., et al. Culex pipiens quinquefasciatus: a potential vector to transmit Zika virus. Emerging Microbes & Infections. 5 (9), 102 (2016).
- Secundino, N. F. C., et al. Zika virus transmission to mouse ear by mosquito bite: a laboratory model that replicates the natural transmission process. Parasites & Vectors. 10 (1), 346 (2017).
- Smith, D. R., et al. Venezuelan Equine Encephalitis Virus Transmission and Effect on Pathogenesis. Emerging Infectious Diseases. 12 (8), 1190-1196 (2006).
- Lazear, H. M., et al. A Mouse Model of Zika Virus Pathogenesis. Cell Host Microbe. 19 (5), 720-730 (2016).
- Morrison, T. E., Diamond, M. S. Animal Models of Zika Virus Infection, Pathogenesis, and Immunity. Journal of Virology. 91 (8), 9-17 (2017).
- Reynolds, E. S., Hart, C. E., Hermance, M. E., Brining, D. L., Thangamani, S. An Overview of Animal Models for Arthropod-Borne Viruses. Comparative Medicine. 67 (3), 232-241 (2017).
- Rossi, S. L., et al. Characterization of a Novel Murine Model to Study Zika Virus. American Journal of Tropical Medicine and Hygiene. 94 (6), 1362-1369 (2016).
- Styer, L. M., et al. Mosquitoes inoculate high doses of West Nile virus as they probe and feed on live hosts. PLoS Pathogens. 3 (9), 1262-1270 (2007).
- Azar, S. R., Diaz-Gonzalez, E. E., Danis-Lonzano, R., Fernandez-Salas, I., Weaver, S. C. Naturally infected Aedes aegypti collected during a Zika virus outbreak have viral titres consistent with transmission. Emerging Microbes & Infections. 8 (1), 242-244 (2019).
- Dzul-Manzanilla, F., et al. Evidence of vertical transmission and co-circulation of chikungunya and dengue viruses in field populations of Aedes aegypti (L.) from Guerrero, Mexico. Transactions of the Royal Society of Tropical Medicine and Hygiene. 110 (2), 141-144 (2016).
- Grard, G., et al. Zika virus in Gabon (Central Africa) – 2007: a new threat from Aedes albopictus. PLoS Neglected Tropical Diseases. 8 (2), 2681 (2014).
- Grubaugh, N. D., et al. Genomic epidemiology reveals multiple introductions of Zika virus into the United States. Nature. 546 (7658), 401-405 (2017).
- Guerbois, M., et al. Outbreak of Zika Virus Infection, Chiapas State, Mexico, 2015, and First Confirmed Transmission by Aedes aegyti Mosquitoes in the America. The Journal of Infectious Diseases. 214 (9), 1349-1356 (2016).
- Lundstrom, J. O., et al. Sindbis virus polyarthritis outbreak signalled by virus prevalence in the mosquito vectors. PLoS Neglected Tropical Diseases. 13 (8), 0007702 (2019).
- Miller, B. R., Monath, T. P., Tabachnik, W. J., Ezike, V. I. Epidemic yellow fever caused by an incompetent mosquito vector. Tropical Medicine and Parasitology. 40 (4), 396-399 (1989).
- Brown, H. E., et al. Effectiveness of Mosquito Traps in Measuring Species Abundance and Composition. Journal of Medical Entomology. 45 (3), 517-521 (2008).
- Gorsich, E. E., et al. A comparative assessment of adult mosquito trapping methods to estimate spatial patterns of abundance and community composition in southern Africa. Parasites & Vectors. 12 (1), 462 (2019).
- Azar, S. R., et al. ZIKV Demonstrates Minimal Pathologic Effects and Mosquito Infectivity in Viremic Cynomolgus Macaques. Viruses. 10 (11), 661 (2018).
- Azar, S. R., et al. Differential Vector Competency of Aedes albopictus Populations from the Americas for Zika Virus. American Journal of Tropical Medicine and Hygiene. 97 (2), 330-339 (2017).
- Hanley, K. A., Azar, S. R., Campos, R. K., Vasilakis, N., Rossi, S. L. Support for the Transmission-Clearance Trade-Off Hypothesis from a Study of Zika Virus Delivered by Mosquito Bite to Mice. Viruses. 11 (11), 1072 (2019).
- Hart, C. E., et al. Zika Virus Vector Competency of Mosquitoes, Gulf Coast, United States. Emerging Infectious Diseases. 23 (3), 559-560 (2017).
- Karna, A. K., et al. Colonized Sabethes cyaneus, a Sylvatic New World Mosquito Species, Shows a Low Vector Competence for Zika Virus Relative to Aedes aegypti. Viruses. 10 (8), 434 (2018).
- Roundy, C. M., et al. Variation in Aedes aegypti Mosquito Competence for Zika Virus Transmission. Emerging Infectious Diseases. 23 (4), 625-632 (2017).
- Wilson, A. J., Harrup, L. E. Reproducibility and relevance in insect-arbovirus infection studies. Current Opinion in Insect Science. 28, 105-112 (2018).
- Hagan, R. W., et al. Dehydration prompts increased activity and blood feeding by mosquitoes. Scientific Reports. 8 (1), 6804 (2018).
- Guo, X. X., et al. Host Feeding Patterns of Mosquitoes in a Rural Malaria-Endemic Region in Hainan Island, China. Journal of the American Mosquito Control Association. 30 (4), 309-311 (2014).
- Kuno, G. Early history of laboratory breeding of Aedes aegypti (Diptera: Culicidae) focusing on the origins and use of selected strains. Journal of Medical Entomology. 47 (6), 957-971 (2010).
- Mayilsamy, M. Extremely Long Viability of Aedes aegypti (Diptera: Culicidae) Eggs Stored Under Normal Room Condition. Journal of Medical Entomology. 56 (3), 878-880 (2019).
- Althouse, B. M., et al. Potential for Zika Virus to Establish a Sylvatic Transmission Cycle in the Americas. PLoS Neglected Tropical Diseases. 10 (12), 0002055 (2016).
- Vasilakis, N., Cardosa, J., Hanley, K. A., Holmes, E. C., Weaver, S. C. Fever from the forest: prospects for the continued emergence of sylvatic dengue virus and its impact on public health. Nature Reviews Microbiology. 9 (7), 532-541 (2011).
- Vasilakis, N., et al. Potential of ancestral sylvatic dengue-2 viruses to re-emerge. Virology. 358 (2), 402-412 (2007).
