For an initial proof of concept experiment, donor endometrium from RFP mice was injected into wildtype recipient mice. H&E staining revealed histopathologic confirmation of classic architecture of endometriosis lesion (Figure 3A). Fluorescent microscopy confirmed that the observed lesion in question originated from the donor (Figure 3B).
The second experiment was performed using 10 wildtype C57BL/6J donors and 10 recipients. An additional 5 recipients received sham treatment (injected with PBS and not endometrial fragments) and assigned a random identification number; reviewers were blinded prior to necropsy and histopathologic review.
All recipients received donor endometrial tissue within 42 h of PSMG donor treatment. There was no correlation between this time interval and final uterine weight or lesion size. At the time of donor procurement, average total uterine weight was 54 ± 9.5 mg. Average fragment number was 22.4 ± 5.2, resulting in average fragment weight of 2.5 ± 0.5 mg. Endpoint surgery occurred at either day 20 or day 22 for all recipients.
With an average number of lesions of 1.5 and average gross total lesion diameter of 3.7 mm, these end points are similar to other studies utilizing mouse models with a similar weight of injected endometrial fragments (Figure 2)10. The prevalence of lesions for all mice was 80%. The mice without lesions had significantly greater endometrial fractioning compared to mice with lesions (30.5 total fragments versus 20.3, respectively; average total fragments for all mice was 22.4) resulting in below average fragment size at the time of injection (1.9 mg vs 2.6 mg). As expected, sham controls (injected with saline and not endometrial fragments) had no gross or microscopic disease. In a subsequent study using an expanded number of mice, estrous phase of the recipient mouse was not associated with lesion number or microscopic disease score.
Gross lesion number appears to be a poor surrogate marker for total lesion burden, as there was discordance between the macroscopic and the microscopic disease present in the tissues. In 60% (n = 6) of the cases, there was agreement between the macroscopic and microscopic scoring (agreement defined as both showing presence or absence and difference in total size was within 3 mm). However, in 40% (n = 4) of the cases, there was disagreement, with 10% (n = 1) of the time macroscopic disease was absent but microscopic disease was present by histology, 10% (n = 1) macroscopic disease was seen despite no absence of endometriosis on confirmatory histology (Figure 5), and the remaining 20% (n = 2) there was agreement in presence but not magnitude of lesion size. Thus, macroscopic examination for lesions alone is not sufficient for the quantification of endometriosis disease burden.
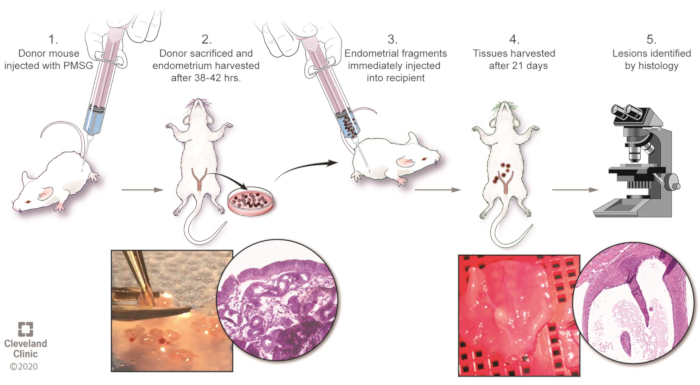
Figure 1: Overview of the model. Please click here to view a larger version of this figure.
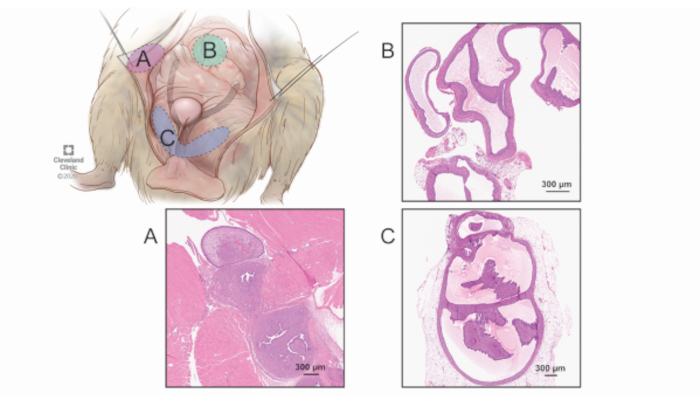
Figure 2: Surveyed regions for lesion quantification. While the bowel and other intraperitoneal locations can harbor lesions, it is more challenging to distinguish these grossly and an exhaustive survey of the abdomen would be more time intensive with diminishing returns. Therefore, a consistent, systematic approach of harvesting the complete tissue (regardless of whether gross disease is present) is performed in the three most common regions of endometriosis formation: the abdominal wall/peritoneum (A), the pancreas and mesenteric fat (B), and the parauterine fat (C). Labeled are representative images of microscopic findings of lesions from each of the three regions. 40x magnification. Please click here to view a larger version of this figure.
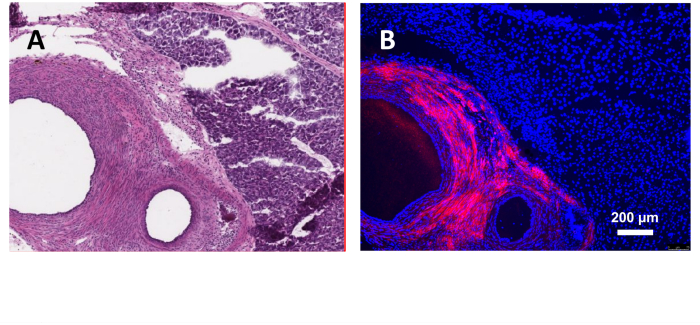
Figure 3: Induction of endometriosis using donor mice endometrium expressing red fluorescent protein. (A) H&E section of the lesion. (B) Fluorescent microscopy with DAPI staining. 40x magnification. Please click here to view a larger version of this figure.
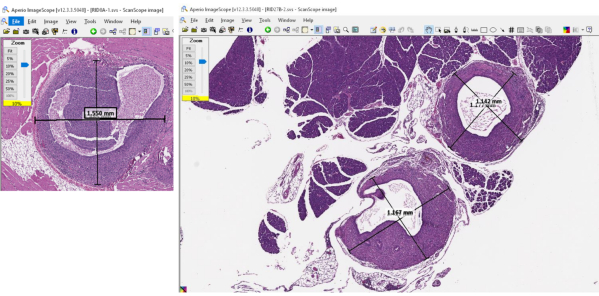
Figure 4: Representative images of the software used to measure the dimensions of the lesion dimensions and quantify lesion burden. Please click here to view a larger version of this figure.
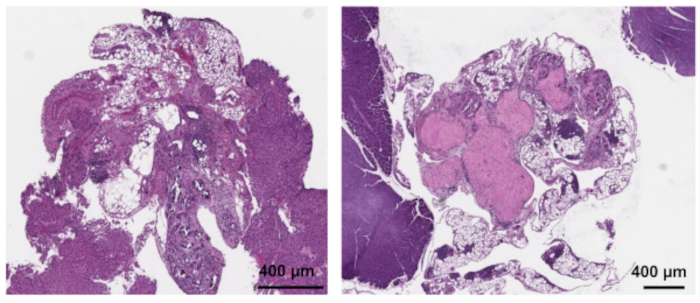
Figure 5: Representative samples of gross "lesions" that are not endometriosis by histopathologic examination. 40x magnification. Please click here to view a larger version of this figure.
